
Sifonòfors
| No s'ha de confondre amb Sifonofòrides. |
| Siphonophorae | |
|---|---|
.jpg) Sifonòfor marí | |
| Enregistrament | |
| Període | |
| Taxonomia | |
| Superregne | Holozoa |
| Regne | Animalia |
| Subregne | Eumetazoa |
| Fílum | Cnidaria |
| Classe | Hydrozoa |
| Ordre | Siphonophorae Eschscholtz, 1829 |
| Nomenclatura | |
| Sinònims | Siphonophora Eschscholtz, 1928
|
| Subordres | |
Els sifonòfors o Siphonophorae (del grec «siphōn», tub + «pherein», portar)[2] és un ordre d'hidrozous, una classe d'organismes marins pertanyents al fílum Cnidaria. Segons el Registre Mundial d'Espècies Marines (World Register of Marine Species, WorMS), l'ordre conté 175 espècies.[3]
Tot i que un sifonòfor pot semblar un organisme individual, cada exemplar és de fet un organisme colonial format per zooides, medusoides i polipoides especialitzats morfològicament i funcionalment.[4] Els zooides són unitats pluricel·lulars que es desenvolupen a partir d'un únic ou fecundat i es combinen per crear colònies funcionals capaces de reproduir-se, digerir, flotar, mantenir la posició corporal i utilitzar la propulsió a raig per moure's.[5] La majoria de colònies són llargues, primes i transparents que viuen surant a la zona pelàgica.[6]
Com altres hidrozous, alguns sifonòfors emeten llum per atreure i atacar les preses. Mentre que molts animals marins produeixen bioluminescència blava i verda, un sifonòfor del gènere Erenna va ser només la segona forma de vida que es va trobar per produir una llum vermella (la primera és el Chirostomias pliopterus).[7][8]
Anatomia i morfologia
[modifica]Característiques de la colònia
[modifica]Els sifonòfors són hidrozous colonials que no presenten alternança de generacions, sinó que es reprodueixen de manera asexual mitjançant un procés de gemmació.[9] Els zooides són les unitats pluricel·lulars que construeixen les colònies. Un únic brot anomenat pro-brot inicia el creixement d'una colònia mitjançant la fissió.[6] Cada zooide es produeix per ser genèticament idèntic; tanmateix, les mutacions poden alterar les seves funcions i augmentar la diversitat dels zooides dins de la colònia.[6] Els sifonòfors són únics perquè el pro-brot inicia la producció de diversos zooides amb funcions específiques.[6] Les funcions i organitzacions dels zooides en colònies varien àmpliament entre les diferents espècies; tanmateix, la majoria de colònies estan disposades bilateralment amb els costats dorsal i ventral de la tija.[6] La tija és la branca vertical al centre de la colònia a la qual s'uneixen els zooides.[6] Els zooides solen tenir funcions especials i, per tant, assumeixen patrons espacials específics al llarg de la tija.[6]
Morfologia general
[modifica]Els sifonòfors solen presentar un dels tres plans corporals estàndard. Els plans corporals s'anomenen Cystonecta, Physonecta i Calycophorae:[10]
- Els «cistonects» tenen una tija llarga amb els zooides units.[10] Cada grup de zooides té un gastrozooide.[10] El gastrozooide té un tentacle utilitzat per capturar i digerir els aliments.[10] Els grups també disposen de gonòfors, especialitzats per a la reproducció.[10] Utilitzen un pneumatòfor, un flotador ple de gas, al seu extrem anterior i principalment es desplacen a la superfície de l'aigua.[10]
- Els «fisonects» tenen un pneumatòfor i un nectosoma, que alberga els nectòfors utilitzats per a la propulsió a raig.[10] Els nèctòfors bombejen l'aigua cap enrere per avançar.[10]
- Els «calicòfors» es diferencien dels citonects i fisonects perquè tenen dos nectòfors i cap pneumatòfor.[10]
Des del seu origen s'ha observat un augment del nombre de tipus zooides en els sifonòfors.[11] Els científics han determinat dues possibles hipòtesis evolutives per a aquesta observació:
- A mesura que ha passat el temps, la quantitat de tipus de zooides ha augmentat.[11]
- L'últim avantpassat comú tenia molts tipus de zooides i la diversitat que es veu avui es deu a la pèrdua de tipus de zooides.[11]
La investigació no mostra cap evidència que recolzi la primera hipòtesi, i s'han vist algunes evidències en suport de la segona.[11]
- Zooides: Els sifonòfors poden tenir zooides que són pòlips o meduses.[12] Tanmateix, els zooides són únics i poden desenvolupar-se per tenir funcions diferents.[12]
- Nectòfors: Els nectòfors són meduses que ajuden a la propulsió i el moviment d'alguns sifonòfors a l'aigua.[5] Són característics en fisonects i calicòfors. Els nectòfors d'aquests organismes es troben al nectosoma on poden coordinar la natació de les colònies.[5] També s'ha observat que els nectòfors treballen conjuntament amb estructures reproductives per tal de proporcionar propulsió durant el despreniment de la colònia.[5]
- Bràctees: Les bràctees són zooides exclusius de l'ordre de les sifonòfors. Funcionen en protecció i mantenint una flotabilitat neutra.[5] Tanmateix, les bràctees no estan presents en totes les espècies de sifonòfors.[5]
- Gastrozooides: Els gastrozooides són pòlips que han desenvolupat una funció per ajudar a l'alimentació dels sifonòfors.[13]
- Palpons: Els palpons són gastrozooides modificats que funcionen en la digestió regulant la circulació dels líquids gastrovasculars.[5]
- Gonòfors :Els gonòfors són zooides que intervenen en els processos reproductius dels sifonòfors.[5]
- Pneumatòfors: La presència de pneumatòfors caracteritza els subgrups Cystonectae i Physonectae.[14] Són flotadors plens de gas que es troben a l'extrem anterior de les colònies d'aquestes espècies.[5] Funcionen per ajudar les colònies a mantenir la seva orientació a l'aigua.[5] En el subgrup Cystonectae, els pneumatòfors tenen una funció addicional d'ajudar a la flotació dels organismes.[5] Els sifonòfors que presenten la característica desenvolupen l'estructura en el desenvolupament primerenc de la larva mitjançant invaginacions de l'estructura de la plànula aplanada.[5] Observacions addicionals de l'espècie de sifonòfor Nanomia bijuga indiquen que la característica del pneumatòfor també funciona potencialment per detectar els canvis de pressió i regular la quimiotaxi en algunes espècies.[15]
Distribució i hàbitat
[modifica]Actualment, el Registre Mundial d'Espècies Marines (WoRMS) identifica 175 espècies de sifonòfors.[10] Poden diferir molt pel que fa a la mida i la forma, cosa que reflecteix en gran manera l'entorn on habiten.[10] Els sifonòfors solen ser organismes pelàgics, però les espècies de nivell són bentòniques.[10] Els sifonòfors d'aigua calenta més petits solen viure a la zona epipelàgica i utilitzen els seus tentacles per capturar zooplàncton i copèpodes.[10] Els sifonòfors més grans viuen en aigües més profundes, ja que generalment són més llargs i fràgils i han d'evitar els forts corrents.[10] S'alimenten principalment de preses més grans. La majoria dels sifonòfors viuen a les profunditats del mar i es poden trobar a tots els oceans.[10] Les espècies de sifonòfors rarament només habiten un lloc.[10] Algunes espècies, però, es poden limitar a un rang específic de profunditats i/o una zona de l'oceà.[10]
Comportament
[modifica]Moviment
[modifica].jpg)
Els sifonòfors utilitzen un mètode de locomoció semblant a la propulsió a raig d'aigua. Un sifonòfor és una colònia agregada complexa formada per molts nectòfors, que són individus clonals que es formen per gemmació i són genèticament idèntics.[16] Segons on es col·loqui cada nectòfor individual dins del sifonòfor, la seva funció és diferent.[16] El moviment colonial està determinat per nectòfors individuals de totes les etapes de desenvolupament. Els individus més petits es concentren cap a la part superior del sifonòfor, i la seva funció és girar i ajustar l'orientació de la colònia.[16] Els individus es faran més grans a mesura que envelleixen. Els individus més grans es troben a la base de la colònia, i la seva funció principal és la propulsió d'empenta.[16] Aquests individus més grans són importants per assolir la velocitat màxima de la colònia.[16] Cada individu és clau per al moviment agregat de la colònia, i entendre la seva organització ens pot permetre avançar en els nostres propis vehicles de propulsió multijet.[16]
L'organització colonial dels sifonòfors, particularment a Nanomia bijuga, confereix avantatges evolutius.[16] Un gran nombre d'individus concentrats permet la redundància.[16] Això vol dir que, fins i tot si alguns nectòfors individuals es veuen compromesos funcionalment, el seu paper es passa per alt, de manera que la colònia en el seu conjunt no es vegi afectada negativament.[16] El velum, una banda fina de teixit que envolta l'obertura del raig d'aigua, també juga un paper en els patrons de natació, mostrats específicament a través de la investigació realitzada sobre l'espècie esmentada anteriorment Namonia bijuga.[17] El velum es fa més petit i circular durant els temps de propulsió cap endavant en comparació amb un velum gran que es veu durant els períodes de recàrrega.[17] A més, la posició del velum canvia amb els comportaments de natació; el velum es corba cap avall en temps de llançament, però durant l'ompliment, el velum es trasllada de nou al nectòfor.[17] El sifonòfor Namonia bijuga també practica la migració vertical diària, ja que roman al fons del mar durant el dia però puja durant la nit.[16]
Depredació i alimentació
[modifica]Els sifonòfors són depredadors carnívors.[4] La seva dieta consisteix en una varietat de copèpodes, petits crustacis i peixos petits.[4] Generalment, les dietes dels sifonòfors nadadors forts consisteixen en preses més petites, i les dietes dels sifonòfors nadadors febles consisteixen en preses més grans.[18]
-
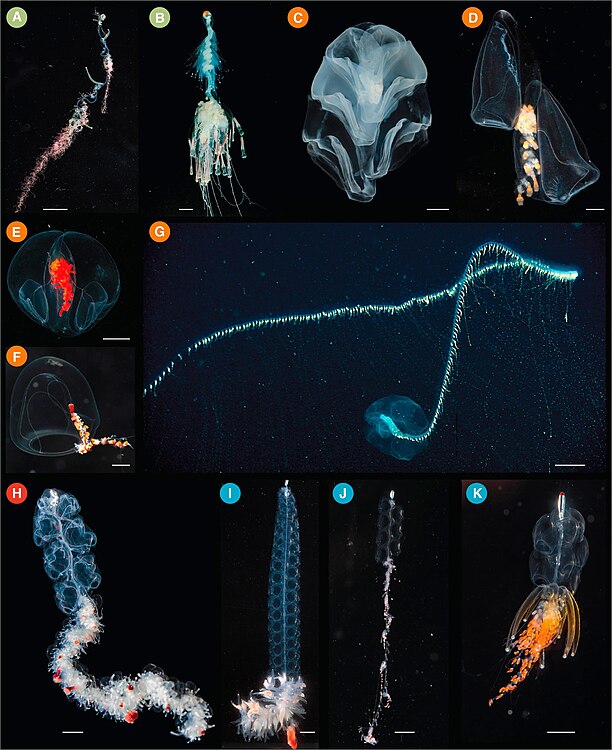 (A) Rhizophysa eysenhardtii, escala de la barra = 1 cm. (B) Bathyphysa conifera, escala de la barra = 2 cm. (C) Hippopodius hippopus, escala de la barra = 5 mm. (D) Kephyes hiulcus, escala de la barra = 2 mm. (E) Desmophyes haematogaster, escala de la barra = 5 mm. (F) Sphaeronectes christiansonae, escala de la barra = 2 mm. (G) Praya dubia, escala de la barra = 4 cm. (H) Apolemia sp., escala de la barra = 1 cm. (I) Lychnagalma utricularia, escala de la barra = 1 cm. (J) Nanomia sp., escala de la barra = 1 cm. (K) Physophora hydrostatica, escala de la barra = 5 mm.
(A) Rhizophysa eysenhardtii, escala de la barra = 1 cm. (B) Bathyphysa conifera, escala de la barra = 2 cm. (C) Hippopodius hippopus, escala de la barra = 5 mm. (D) Kephyes hiulcus, escala de la barra = 2 mm. (E) Desmophyes haematogaster, escala de la barra = 5 mm. (F) Sphaeronectes christiansonae, escala de la barra = 2 mm. (G) Praya dubia, escala de la barra = 4 cm. (H) Apolemia sp., escala de la barra = 1 cm. (I) Lychnagalma utricularia, escala de la barra = 1 cm. (J) Nanomia sp., escala de la barra = 1 cm. (K) Physophora hydrostatica, escala de la barra = 5 mm.

La majoria dels sifonòfors tenen gastrozooides que tenen un tentacle característic unit a la base del zooide. Aquesta característica estructural funciona per ajudar els organismes a capturar preses.[5] Les espècies amb grans gastrozooides són capaços de consumir una àmplia gamma de mides de preses.[18] De manera similar a molts altres organismes del fílum de Cnidaria, moltes espècies de sifonòfors presenten càpsules urticants de nematocists a les branques dels seus tentacles anomenades tentilla.[5] Els nematocists estan disposats en bateries denses al costat de les tentilla.[5] Quan el sifonòfor es troba amb preses potencials, el seu tentillum reacciona on els tentacles de 30 a 50 cm creen una xarxa transformant la seva forma al voltant de la presa.[4][19][20] Aleshores, els nematocists disparen milions de molècules de toxines paralitzants,[18] i de vegades mortals a la presa atrapada que després es trasllada al lloc adequat per a la digestió.[4] Algunes espècies de sifonòfors utilitzen un mimetisme agressiu mitjançant l'ús de llum bioluminescent perquè la presa no pugui identificar correctament el depredador.[19]
Hi ha quatre tipus de nematoquists a les tentilla del sifonòfor: heteronemes, haplonemes, desmonemes i ropalonemes.[19]
- Els heteronemes són els nematoquists més grans i són espines en un eix proper als túbuls units al centre del sifonòfor.[19]
- Els haplonemes tenen túbuls amb punta oberta amb espines, però sense un eix diferenciat.[19] Aquest és el nematoquist més comú entre els sifonòfors.[19]
- Els desmonemes no tenen espines, sinó que hi ha propietats adhesives als túbuls per subjectar-se a les preses.[19]
- Els ropalonemes són nematoquists amb túbuls amples per a les preses.[19]
A causa de la manca d'aliment a l'entorn de les profunditats marines, la majoria d'espècies de sifonòfors funcionen en una tàctica d'estar quiets i esperar que arribi el menjar.[21] El pla corporal gelatinós permet flexibilitat a l'hora de capturar preses, però les adaptacions gelatinoses es basen en l'hàbitat.[22] Neden tot esperant que els seus llargs tentacles es trobin amb les preses. A més, els sifonòfors d'un grup denominat Erenna tenen la capacitat de generar bioluminescència i fluorescència vermella mentre les seves tentilla es contrauen de tal manera que imita els moviments de petits crustacis i copèpodes.[8] Aquestes accions atrauen a la presa perquè s'apropi al sifonòfor, permetent-lo atrapar i digerir-lo.[8]
Reproducció
[modifica]Els modes de reproducció dels sifonòfors varien entre les diferents espècies i, fins avui, es desconeixen diversos modes. Generalment, un sol zigot comença la formació d'una colònia de zooides.[4] L'òvul fecundat madura en un protozooide, que inicia el procés de germinació i la creació d'un nou zooide.[4] Aquest procés es repeteix fins que es forma una colònia de zooides al voltant de la tija central.[4] En canvi, diverses espècies es reprodueixen mitjançant pòlips. Els pòlips poden contenir òvuls i/o espermatozoides i es poden alliberar a l'aigua des de l'extrem posterior del sifonòfor.[4] Els pòlips poden ser fecundats fora de l'organisme.[4]
Els sifonòfors utilitzen gonòfors per fer els gàmetes reproductors.[13] Els gonòfors són masculins o femenins; tanmateix, els tipus de gonòfors d'una colònia poden variar segons les espècies.[13] Les espècies es caracteritzen com a monoiques o dioiques en funció dels seus gonòfors.[13] Les espècies monoiques contenen gonòfors masculins i femenins en una sola colònia zooide, mentre que les espècies dioiques alberguen gonòfors masculins i femenins per separat en diferents colònies de zooides.[13]
Bioluminescència
[modifica]
Gairebé tots els sifonòfors tenen capacitats bioluminescents. Com que aquests organismes són extremadament fràgils, poques vegades s'observen vius.[8] S'ha pensat que la bioluminescència en sifonòfors ha evolucionat com a mecanisme de defensa.[8] Es creu que els sifonòfors del gènere d'aigües profundes Erenna (que es troben a profunditats d'entre 1.600 i 2.300 metres) també utilitzen la seva capacitat bioluminiscent per alimentar-se, com a esquer per atraure peixos.[8] Aquest gènere és un dels pocs que depreda peixos en lloc de crustacis.[8] Els òrgans bioluminescents, anomenats tentilla, d'aquests individus no visuals emeten fluorescència vermella juntament amb un patró de parpelleig rítmic, que atreu les preses ja que s'assembla a organismes més petits com el zooplàncton i els copèpodes. Així, s'ha conclòs que utilitzen la luminescència com a esquer per atreure les preses.[8] Algunes investigacions indiquen que els organismes de les profunditats marines no poden detectar longituds d'ona llargues i la llum vermella té una longitud d'ona llarga de 680 nm. Si aquest és el cas, aleshores els peixos no són atrets per Erenna i hi ha d'haver una altra explicació. No obstant això, les aigües profundes romanen en gran part inexplorades i la sensibilitat a la llum vermella en peixos com Cyclothone i els peixos mictòfids profunds no s'han de descartar.[8]
Els esquers bioluminiscents es troben en moltes espècies diferents de sifonòfors, i s'utilitzen per diverses raons. Espècies com Agalma okeni, Athorybia rosacea, Athorybia lucida i Lychnafalma utricularia utilitzen els seus esquers com a dispositiu de mimetisme per atreure preses.[10] Es creu que A. rosacea imiten les larves de peixos, A. lucida imiten els refugis de larvacis, i L. utricularia imita la hidromedusa.[10] L'espècie Resomia ornicephala utilitza la seva tentilla fluorescent verda i blava per atreure el krill, ajudant-la a superar altres organismes que estan caçant la mateixa presa.[10] Els sifonòfors del gènere Erenna utilitzen esquers bioluminescents envoltats de fluorescència vermella per atreure preses, i possiblement imiten un peix del gènere Cyclothone.[10] Les seves preses són atretes a través d'un comportament únic associat a la tentilla.[8] Quan és jove, la tentilla dels organismes del gènere Erenna només conté teixit bioluminescent, però, a mesura que l'organisme envelleix, també hi ha material fluorescent vermell en aquests teixits.[8]
Taxonomia
[modifica]Els organismes de l'ordre dels sifonòfors s'han classificat en el fílum Cnidaria i la classe Hydrozoa.[3] Les relacions filogenètiques dels sifonòfors han estat de gran interès per l'alta variabilitat de l'organització de les seves colònies de pòlips i meduses.[23][13] Abans es creia que era un grup molt diferent, les similituds larvàries i les característiques morfològiques han fet creure als investigadors que els sifonòfors havien evolucionat a partir d'hidrozous colonials més simples similars als dels ordres Anthoathecata i Leptothecata.[14] En conseqüència, ara s'uneixen amb aquests a la subclasse Hydroidolina.
Les primeres anàlisis van dividir els sifonòfors en 3 subgrups principals en funció de la presència o absència de dos trets diferents: campanes nedadores (nectòfors) i flotadors (neumatòfors).[14] Els subgrups estaven formats per Cystonectae, Physonectae i Calycorphores. Cystonectae tenia pneumatòfors, Physonectae tenien nèctòfors, i Calycorphores tenia tots dos.[14]
El gen ribosòmic de la subunitat petita nuclear eucariota 18S, el gen ribosòmic de la subunitat gran mitocondrial eucariota 16S, i les anàlisis del transcriptoma avalen encara més la divisió filogenètica de Siphonophorae en dos clades principals: Cystonectae i Codonophora. Els subordres de Codonophora inclouen Physonectae (constituïts pels clades Calycophorae i Euphysonectae), Pyrostephidae i Apolemiidae.[5][13]
- Subordre Calycophorae
- Abylidae Agassiz, 1862
- Clausophyidae Totton, 1965
- Diphyidae Quoy & Gaimard, 1827
- Hippopodiidae Kölliker, 1853
- Prayidae Kölliker, 1853
- Sphaeronectidae Huxley, 1859
- Tottonophyidae Pugh, Dunn & Haddock, 2018
- Subordre Cystonectae
- Physaliidae Brandt, 1835
- Rhizophysidae Brandt, 1835
- Subordre Physonectae
- Agalmatidae Brandt, 1834
- Apolemiidae Huxley, 1859
- Cordagalmatidae Pugh, 2016
- Erennidae Pugh, 2001
- Forskaliidae Haeckel, 1888
- Physophoridae Eschscholtz, 1829
- Pyrostephidae Moser, 1925
- Resomiidae Pugh, 2006
- Rhodaliidae Haeckel, 1888
- Stephanomiidae Huxley, 1859
Història
[modifica]Descobriment
[modifica]Carl Linnaeus va descobrir i descriure el primer sifonòfor, la caravel·la portuguesa (Physalia physalis), l'any 1758.[10]
La taxa de descobriment d'espècies de sifonòfors va ser lenta al segle xviii, ja que només es van trobar quatre espècies addicionals.[10]
Durant el segle xix es van observar 56 espècies noves a causa dels viatges de recerca realitzats per les potències europees.[10] La majoria de les espècies noves trobades durant aquest període de temps es van recollir a les aigües costaneres i superficials.[10] Durant l'expedició del HMS Challenger, es van recollir diverses espècies de sifonòfors. Ernst Haeckel va intentar fer un escrit de totes les espècies de sifonòfors recollides en aquesta expedició. Va introduir 46 «espècies noves»; tanmateix, el seu treball va ser molt criticat perquè finalment es va trobar que algunes de les espècies que va identificar no eren sifonòfors.[10] No obstant això, algunes de les seves descripcions i figures (imatges del següent apartat) són considerades útils pels biòlegs moderns.
Durant el segle xx es va observar una taxa d'uns 10 descobriments d'espècies noves per dècada.[10] Considerat l'investigador més important dels sifonòfors, A. K. Totton va introduir 23 noves espècies de sifonòfors a mitjans del segle xx.[10]
El 6 d'abril de 2020, el Schmidt Ocean Institute va anunciar el descobriment d'un sifonòfor gegant, Apolemia, als canyons submarins prop de la costa de Ningaloo, de 15 m de diàmetre amb un anell d'aproximadament 47 m de llarg, possiblement el sifonòfor més gran mai gravat.[24][25]
No hi ha registre fòssil de sifonòfors, tot i que han evolucionat i s'han adaptat durant un llarg període de temps. El seu fílum, Cnidaria, és un antic llinatge que es remunta al c. fa 640 milions d'anys.[10]
Sifonòfors de Haeckel
[modifica]Ernst Haeckel va descriure una sèrie de sifonòfors, i diverses làmines del seu Kunstformen der Natur (1904) representen membres del tàxon:[26]
-
 Làmina 7
Làmina 7 -
 Làmina 37
Làmina 37 -
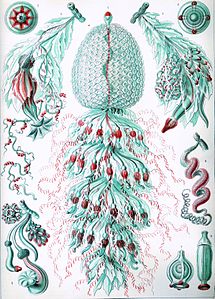 Làmina 59
Làmina 59 -
 Làmina 77
Làmina 77




Referències
[modifica]- ↑ Entrada «Siphonophora» de la Paleobiology Database (en anglès). [Consulta: 20 desembre 2022].
- ↑ «Siphonophora» (en anglès). Lexico. Arxivat de l'original el 2020-10-22. [Consulta: 26 agost 2023].
- ↑ 3,0 3,1 «"Siphonophorae"» (en anglès). World Register of Marine Species (WoRMS), 2018.
- ↑ 4,00 4,01 4,02 4,03 4,04 4,05 4,06 4,07 4,08 4,09 «Pelagic Siphonophore» (en anglès). Aquarium of Pacific.
- ↑ 5,00 5,01 5,02 5,03 5,04 5,05 5,06 5,07 5,08 5,09 5,10 5,11 5,12 5,13 5,14 5,15 Munr, Catriona; Siebert, Stefan; Zapata, Felipe; Howison, Mark; et al. «Improved phylogenetic resolution within Siphonophora (Cnidaria) with implications for trait evolution» (en anglès). bioRxiv, 20-01-2018. DOI: 10.1101/251116.
- ↑ 6,0 6,1 6,2 6,3 6,4 6,5 6,6 Dunn, 2005, p. 835-845.
- ↑ «Siphonophores» (en anglès). Smithsonian Ocean.
- ↑ 8,00 8,01 8,02 8,03 8,04 8,05 8,06 8,07 8,08 8,09 8,10 Haddok et al., 2005, p. 263.
- ↑ Pugh, 2014.
- ↑ 10,00 10,01 10,02 10,03 10,04 10,05 10,06 10,07 10,08 10,09 10,10 10,11 10,12 10,13 10,14 10,15 10,16 10,17 10,18 10,19 10,20 10,21 10,22 10,23 10,24 10,25 10,26 10,27 10,28 Mapstone, Gillian M «Global Diversity and Review of Siphonophorae (Cnidaria: Hydrozoa)» (en anglès). PLOS One, 9(2), 06-02-2014, pàg. e87737. Bibcode: 2014PLoSO...987737M. DOI: 10.1371/journal.pone.0087737. ISSN: 1932-6203. PMC: 3916360. PMID: 24516560.
- ↑ 11,0 11,1 11,2 11,3 Dunn, Casey W; Pugh, Philip R; Haddock, Steven H. D «Molecular Phylogenetics of the Siphonophora (Cnidaria), with Implications for the Evolution of Functional Specialization» (en anglès). Systematic Biology, 54(6), 01-12-2005, pàg. 916-935. DOI: 10.1080/10635150500354837. ISSN: 1076-836X. PMID: 16338764.
- ↑ 12,0 12,1 Dunn, 2009, p. R233-R234.
- ↑ 13,0 13,1 13,2 13,3 13,4 13,5 13,6 Dunn, Casey W; Pugh, Philip R; Haddock, Steven H. D «Molecular Phylogenetics of the Siphonophora (Cnidaria), with Implications for the Evolution of Functional Specialization» (en anglès). Systematic Biology, 54(6), 01-12-2005, pàg. 916-935. DOI: 10.1080/10635150500354837. ISSN: 1076-836X. PMID: 16338764.
- ↑ 14,0 14,1 14,2 14,3 Collins, Allen G «Phylogeny of Medusozoa and the evolution of cnidarian life cycles» (en anglès). Journal of Evolutionary Biology, 15(3), 30-04-2002, pàg. 418-432. DOI: 10.1046/j.1420-9101.2002.00403.x.
- ↑ Church et al., 2015, p. 435-449.
- ↑ 16,00 16,01 16,02 16,03 16,04 16,05 16,06 16,07 16,08 16,09 Costello et al., Sutherland, p. 8158.
- ↑ 17,0 17,1 17,2 Sutherland, Kelly R; Gemmell, Brad J; Colin, Sean P; Costello, John H «Propulsive design principles in a multi-jet siphonophore» (en anglès). The Journal of Experimental Biology, 222(6), 15-03-2019, pàg. jeb198242. DOI: 10.1242/jeb.198242. ISSN: 0022-0949. PMID: 30814298.
- ↑ 18,0 18,1 18,2 Purcell, 1990, p. 1045-1047.
- ↑ 19,0 19,1 19,2 19,3 19,4 19,5 19,6 19,7 Damian Serrano, Alejandro; Haddock, Steven H.D; Dunn, Casey W «Shaped to kill: The evolution of siphonophore tentilla for specialized prey capture in the open ocean» (en anglès). bioRxiv, 12-06-2019. DOI: 10.1101/653345.
- ↑ Damian Serrano, Alejandro; Haddock, Steven H. D; Dunn, Casey W «Shaped to kill: The evolution of siphonophore tentilla for specialized prey capture in the open ocean» (en anglès). bioRxiv, 02-04-2020, pàg. 653345. DOI: 10.1101/653345.
- ↑ Dunn, Casey. «"Siphonophores"» (en anglès), 2005.
- ↑ Madinand i Harbison, 2001.
- ↑ Waggoner, Ben. «Hydrozoa: More on Morphology» (en anglès). UC Museum of Paleontology, 21-07-1995.
- ↑ «Longest Giant Stringy Sea Creature Ever Recorded Looks like It Belongs in Outer Space» (en anglès). Interesting Engineering, 09-04-2020.
- ↑ Schmidt Ocean Institute. «New species discovered during exploration of abyssal deep sea canyons off Ningaloo» (en anglès). EurekAlert!, 09-04-2020.
- ↑ Costantino, Grace. «Art Forms in Nature: Marine Species From Ernst Haeckel» (en anglès). Smithsonian Ocean (Smithsonian Institution ).
Bibliografia
[modifica]- Church, Samuel H; Siebert, Stefan; Bhattacharyya, Pathikrit; Dunn, Casey W «The histology of Nanomia bijuga (Hydrozoa: Siphonophora)» (en anglès). Journal of Experimental Zoology Part B: Molecular and Developmental Evolution, 324(5), juliol 2015. DOI: 10.1002/jez.b.22629. PMC: 5032985. PMID: 26036693.
- Costello, John H; Colin, Sean P; Gemmell, Brad J; Dabiri, John O; Sutherland, Kelly R «Multi-jet propulsion organized by clonal development in a colonial siphonophore» (en anglès). Nature Communications, 6(1), novembre 2015. Bibcode: 2015NatCo...6.8158C. DOI: 10.1038/ncomms9158. ISSN: 2041-1723. PMC: 4569723. PMID: 26327286.
- Dunn, Casey W «Complex colony-level organization of the deep-sea siphonophore Bargmannia elongata (Cnidaria, Hydrozoa) is directionally asymmetric and arises by the subdivision of pro-buds» (en anglès). Developmental Dynamics, 234(4), desembre 2005. DOI: 10.1002/dvdy.20483. PMID: 15986453.
- Dunn, Casey «Siphonophores» (en anglès). Current Biology, 19(6), 2009, pàg. R233-R234. DOI: 10.1016/j.cub.2009.02.009. PMID: 19321136.
- Haddock, S. H; Dunn, C. W; Pugh, P. R; Schnitzler, C. E «Bioluminescent and red-fluorescent lures in a deep-sea siphonophore» (en anglès). Science, 309(5732), juliol 2005. DOI: 10.1126/science.1110441. PMID: 16002609.
- Madinand, L. P; Harbison, G. R. «Gelatinous Zooplankton*». A: Encyclopedia of Ocean Sciences (en anglès). Oxford: Academic Press, 2001, p. 9-19. DOI 10.1016/b978-012374473-9.00198-3. ISBN 978-0-12-374473-9.
- Mapstone, Gillian M. Siphonophora (Cnidaria, Hydrozoa) of Canadian Pacific waters (en anglès). Ottawa: NRC Research Press, 2009. ISBN 978-0-660-19843-9.
- Pugh, Philip R «Siphonophora» (en anglès). Access Science, 2014. DOI: 10.1036/1097-8542.625800.
- Purcell, Jennifer E «Influence of Siphonophore Behavior upon Their Natural Diets: Evidence for Aggressive Mimicry» (en anglès). Science, 209, 1980. DOI: 10.1126/science.209.4460.1045.
Enllaços externs
[modifica]- «Siphonophore: Deep-sea superorganism» (en anglès). Pink Tentacle, 2008.
- «Tauchen in Norwegen - Kvasefjord (Scubamedia)». YouTube, 30-08-2013.
- «Siphonophore. Pinktentacle3» (en anglès). YouTube, 22-12-2008.
- «Stunning Siphonophore Sighting» (en anglès). Nautilus Live: Explore the ocean LIVE with Dr. Robert Ballard and the Corps of Exploration (Ocean Exploration Trust), 27-06-2014.
- «Deep sea siphonophore». YouTube, 10-04-2017. Imaged by the NOAA Okeanos Explorer on March 14, 2017 at 1,560 meters west of Winslow Reef complex.