
Usuari:Vallue/Animal communication
_great-egret-8243-web_(324180866).jpg)
La comunicació animal és la transferència d’informació d’un o d’un grup d’animals (emissor o emissors) a un o més altres animals (receptor o receptors) que afecta el comportament actual o futur dels animals que la reben. La informació es pot enviar de manera intencionada, com en una exhibició d'emparellament, o sense voler, com en la transferència de l’aroma del depredador a la presa. Aquesta informació es pot transferir a una "audiència" de diversos receptors. La comunicació animal és una àrea d’estudi que creix ràpidament, incorporant disciplines que inclouen l'etologia, la sociologia, la neurologia i la cognició animal . Molts aspectes del comportament animal, com ara l’ús simbòlic del nom, l’expressió emocional, l’aprenentatge i la conducta sexual, s’estan estudiant amb nous punts de vista.
Quan la informació de l’emissor canvia el comportament d’un receptor, la informació es coneix com a “senyal”. La teoria de senyals preveu que per mantenir un senyal a la població, tant l’emissor com el receptor haurien de rebre normalment algun benefici en la interacció. La producció de senyals per part dels remitents, i la percepció i la resposta posterior dels receptors fa pensar que coevolucionaran. [1] Els senyals solen implicar múltiples mecanismes, per exemple, tant visuals com auditius. Perquè un senyal s'entengui, el comportament coordinat tant de l'emissor com del receptor requereix un estudi atent.
Modes
[modifica]
Visual
[modifica]- Gestos : la forma de comunicació més coneguda consisteix en la visualització de parts del cos distintives o moviments corporals distintius. Sovint es produeixen en combinació, de manera que un moviment actua per revelar o emfatitzar una part del cos. Un exemple significatiu és la presentació del progenitor del gavià argentat als seus pollets com a senyal d'alimentació. Com moltes gavines, el gavià argentat té un colors viu als ulls, de color groc amb una taca vermella a la mandíbula inferior a prop de la punta. Quan el progenitor torna al niu amb menjar, es posa sobre el pollet i li posa la presa a terra. Això provoca una resposta de súplica d'un pollet famolenc (picotejant a la taca vermella), que estimula el progenitor a regurgitar els aliments. El senyal complet comporta, doncs, una característica morfològica distintiva (part del cos), la nota de punt vermell; i un moviment distintiu (que s’apropa cap al terra) que fa que la taca vermella sigui molt visible per al pollet. Si bé tots els primats utilitzen alguna forma de gest, [2] Frans de Waal va concloure que els simis i els humans són únics, ja que només utilitzen gestos intencionats per comunicar-se. Va provar la hipòtesi que els gestos evolucionen cap al llenguatge estudiant la conducta gestual de bonobos i ximpanzés .

- Expressió facial : Els gestos facials tenen un paper important en la comunicació animal. Sovint un gest facial és un senyal d’emoció . Els gossos, per exemple, expressen la ira grunyint i mostrant-se les dents. Les orelles s'erigeixen com a mostra d'alarma i s’enfosqueixen, mentre que exposen les dents i fixen la mirada. Jeffrey Mogil va estudiar les expressions facials dels ratolins durant increments del dolor; hi havia cinc expressions facials reconeixibles: estrenyiment orbital, molèstia del nas i galtes, i canvis en l'oïda i els bigotis. [3]
- Mirada (seguiment) : Els animals socials coordinen la seva comunicació mitjançant el seu control de l'orientació amb el cap i els ulls. Aquest comportament ha estat reconegut des de fa temps com un component important de la comunicació durant el desenvolupament humà, i l'estudi de la mirada en els animals s'ha estudiat força recentment. S'han realitzat investigacions amb simis, [4] micos, gossos, [5] ocells, [6] llops i tortugues, i s'han centrat en dues tasques diferents: "seguir la mirada d'un altre a l'espai llunyà" i "seguir [ment] de la mirada d'un altre geomètricament al voltant d'una barrera visual, per exemple, reposicionant-se a si mateix per seguir una mirada quan es troba davant d'una barrera que bloqueja la seva vista ". La primera habilitat s'ha trobat entre una àmplia gamma d'animals, mentre que la segona només s'ha demostrat per als simis, gossos, llops i còrvids (corbs); Els intents de demostrar aquest "miratge geomètric" a titis i Ibis van donar resultats negatius. Els investigadors encara no tenen una imatge clara de la base cognitiva del seguiment de la mirada, però les evidències del desenvolupament indiquen que la mirada "simple" que segueix i la mirada "geomètrica" probablement es basa en diferents mecanismes cognitius. [7]
- Canvi de color: El canvi de color es pot dividir en canvis que es produeixen durant el creixement i desenvolupament, i els que es desencadenen per l’estat d’ànim, el context social o factors abiòtics com la temperatura. Aquests últims es veuen en molts tàxons. Alguns cefalòpodes, com el polp i la sípia, tenen cèl·lules especialitzades de la pell ( cromatòfors) que poden canviar el color aparent, l'opacitat i la reflectivitat de la seva pell. [8] A més del seu ús per al camuflatge, s'utilitzen canvis ràpids en el color de la pell durant la caça i en els rituals d'aparellament. [9] La sípia pot mostrar dos senyals completament diferents alhora en ambdós costats del cos. Quan una sípia mascle corteja una femella davant d'altres mascles, mostra un patró masculí mirant cap a la femella i un model femení mirant cap a fora, per enganyar a altres mascles. Alguns senyals de color es produeixen en cicles. Per exemple, quan un babuí femella comença a ovular, la seva zona anogenital s’infla i es torna vermell/rosa brillant. Això indica als mascles que està disposada a aparellarr-se. [10]
- Comunicació bioluminescent : La comunicació mitjançant la producció de llum es produeix habitualment en els vertebrats i els invertebrats dels oceans, particularment a les profunditats (per exemple, els peixos lofiformes, majoritàriament abissals). Dues formes molt conegudes de bioluminescència terrestre es donen en els lampírids i cucs luminiscents . Altres insectes, larves d’ insectes, anèl·lids, aràcnids i fins i tot espècies de fongs tenen capacitats bioluminescents. Alguns animals bioluminescents produeixen la llum ells mateixos, mentre que d’altres tenen una relació simbiòtica amb els bacteris bioluminescents.
Auditiu
[modifica]

| Crows flocking |
|
|
| A flock of American crows. |
Molts animals es comuniquen mitjançant vocalització. La comunicació vocal serveix per a molts propòsits, incloent-hi rituals d’aparellament, senyals d’avís, transmissió de l'ubicació de fonts d’alimentació i d'aprenentatge social. En diverses espècies, els mascles realitzen sons durant els rituals d’aparellament com una forma de competència contra altres mascles, i com un senyal de la seva presència per les femelles. Entre algunes espècies cal destacar granotes, ratpenats amb cap de martell, cérvols, balenes geperudes, elefants marins i ocells cantadors. [12] Altres casos de comunicació vocal inclouen els senyals d'alarma del mico de Campbell, [13] les senyals territorials de gibons, i l’ús de freqüència en ratpenats de llança gros per distingir-se entre els grups. [14] El mico Vervet realitza un senyal d'alarma diferent per a cadascun dels seus quatre predadors, i les reaccions d'altres micos varien adequadament segons aquest tipus d'avís. Per exemple, si un senyal d'alarma indica la presència d'una pitó, els micos pugen als arbres, mentre que l'alarma "l'àguila" fa que els micos cerquin un amagatall a terra. Els gossets de les praderies també utilitzen senyals complexes que indiquen diferències de depredadors. Segons Con Slobodchikoff i altres investigadors, els senyals dels gossets de praderies comuniquen el tipus, la mida i la velocitat d’un depredador que s’acosta. [15] [16] [17] També s'ha comprovat que el cant de les balenes tenen diferents dialectes segons la regió que habiten. [18]
No tots els animals utilitzen la vocalització com a mitjà de comunicació auditiva. Molts artròpodes freguen parts especials del cos per produir sons. Això es coneix com a estridulació. Els grills i els saltamartins se'ls coneix molt per aquest comportament. Moltes altres espècies utilitzen també la estridulació, incloent crustacis, aranyes, escorpins, vespes, formigues, escarabats, papallones, arnes, milipedes (milpeus) i centèpsids (centpeus) . Un altre mitjà de comunicació auditiva és la vibració de les bufes natatòres en peixos ossis. L’estructura de les bufes nataòries i els seus músculs sonors adjunts varia molt entre les famílies de peixos ossis, donant lloc a una gran varietat de sons. Les parts del cos que impactent entre si, també poden produir senyals auditius. Un exemple d'aqeust senyal és la vibració de la punta de la cua de les serps de cascabell com a senyal d’avís. Altres espècies inclouen el desplegament del picoteig en els ocells, la batuda d'ales dels manaquís ales com a mostra de festeig, i el colpejament al pit en els goril·les . [19]
Olfactiu
[modifica]Tot i ser el mètode més antic de comunicació, la comunicació química és una de les formes menys estudides degut, en part, a la gran abundància de productes químics al nostre entorn i a la dificultat de detectar i mesurar tots els productes químics en una mostra. [19] La capacitat de detectar productes químics en el medi ambient compleix moltes funcions. Es crucial en la detecció d'aliments. Aquesta funció, va sorgir per primera vegada en organismes unicel·lulars ( bacteris ) que vivien als oceans durant els primers moments de l'esclat de la vida a la Terra. A mesura que aquesta funció evolucionava, els organismes van començar a diferenciar entre compostos químics que provenen de recursos, interespecífics (espècies idèntiques, és a dir, parelles i parents) i heterospecífics (espècies diferents, és a dir, competidors i depredadors). Per exemple, la petita espècie de barb roig pot evitar els hàbitats amb una concentració detectable de productes químics associats a una espècie depredadora com el lluç de riu. [20] Les seves cries, amb la capacitat de percebre la presència de depredadors abans que siguin a prop per ser vistes, i que després responguin amb un comportament adaptatiu (com ocultar-se) tenen més probabilitats de sobreviure i reproduir-se. El marcatge de territorialitat i l'olor produit en el marcatge per fregament són formes habituals de comunicació olfactiva en mamífers. [21] [22]
Elèctric
[modifica]L’electrocomunicació també és una forma rara de comunicació entre els animals. S'observa principalment en animals aquàtics, tot i que alguns mamífers terrestres, en particular els ornitorrincss i els equidnes, emeten camps elèctrics que poden ser utilitzats per la comunicació. [23]
Els peixos elèctrics proporcionen un exemple d’electrocomunicació juntament amb l’ electrorecepció . Aquests peixos utilitzen un òrgan elèctric per generar un camp elèctric, que es detecta mitjançant electroreceptors. Les diferències en la forma d'ona electromagnètica i els canvis en el camp transmeten informació sobre espècies, sexe i identitat. Aquests senyals elèctrics es poden generar en resposta a les hormones, als ritmes circadians i a les interaccions amb altres peixos. Alguns depredadors, com els taurons i els rajades, són capaços d'escoltar aquests peixos electrogènics mitjançant electrorecepció passiva.
Tàctil
[modifica]- Per obtenir més informació sobre el mecanisme de tacte, vegeu Sistema somatosensorial i Mecanoreceptors
El tacte és un factor clau en moltes interaccions socials. Aquests són alguns exemples:
- Lluita: en una lluita, el tacte pot ser utilitzat per desafiar un oponent i coordinar els moviments durant la lluita. El perdedor també l’utilitza per indicar la seva submissió.
- L’aparellament: els mamífers sovint inicien l’aparellament per la relació diària en mateix grup, l’acaronant o fregant-se els uns amb els altres. Això proporciona l’oportunitat d’impregnar senyals químics i de valorar els excretats per la parella potencial. El tacte també pot anunciar la intenció del mascle de copular amb la femella, com quan un cangur mascle agafa la cua d’una femella. Durant l’aparellament, els estímuls tàctils són importants pel posicionament de parella, la coordinació i l’estimulació genital.
- Integració social: el tacte s’utilitza àmpliament per a la integració social, i es caracteritza per la socialització d’un animal per part d'un altre. La interacció social té diverses funcions; elimina els paràsits i elos excrements de l’animal cuidat, reafirma el vincle social o la relació jeràrquica entre els animals i dóna a l’individu guia l’oportunitat d’examinar indicis olfactius sobre l’individu cuidat, i afegint-ne d’altres. Aquest comportament s’ha observat en la eusocialitat dels insectes, aus i mamífers.
- Cerca d'aliment: Algunes espècies de formigues recluten companys de treball per a cercar noves fonts d'alimentació. Primer tocant-los amb les antenes i les extremitats anteriors, per després conduir-los a la font d'aliments mantenint el contacte físic. Un altre exemple d'això és la dansa de les abelles. [24]
- Termoregulació: El contacte físic prolongat o l’ amuntegament també serveix per a la integració social. L'agrupament dels individus afavoreix l'intercanvi de calor, juntament amb la transferència d'informació olfactiva o tàctil. Alguns organismes viuen en constant contacte en una colònia, per exemple les colònies de coralls. Quan els individus estan estretament lligats d’aquesta manera, una colònia sencera pot reaccionar als moviments adversos o d’alarma que fan només uns quants individus. En alguns herbivors, les nimfes i larves, els gups d'individus on hi ha contacte perllongat tenen un paper important en la coordinació del grup. Aquestes agrupacions poden prendre la forma d’una processó o una roseta.
Sísmic
[modifica]La comunicació sísmica és l’intercanvi d’informació mitjançant senyals vibracionals generats automàticament i que es transmeten a través d’un substrat com el sòl, l’aigua, les teranyines, les tiges de la planta o una fulla d’herba. Aquesta forma de comunicació presenta diversos avantatges. Per exemple, pot enviar-se independentment dels nivells de llum i soroll, i sol tenir un rang i una persistència curta, cosa que pot reduir el perill de detecció dels depredadors. L’ús de la comunicació sísmica es troba en molts tàxons, com granotes, rates cangur, farumfers, abelles, cucs nematodes, entre altres. Els tetràpodes solen produir ones sísmiques fent un tambor a terra amb una part del cos, un senyal que es detecta pel sàcul del receptor. El saclus és un òrgan de l'orella interna que conté un sac membranós que s'utilitza per a l'equilibri, però també pot detectar ones sísmiques en animals que utilitzen aquesta forma de comunicació. Les vibracions també es poden combinar amb altres tipus de comunicació. [25]
Tèrmic
[modifica]
Diversos serps tenen la capacitat de detectar radiació tèrmica per infrarojos (IR), cosa que permet que aquests rèptils puguin obtenir imatges tèrmiques de la calor radiant emesa pels depredadors o preses a longituds d'ona entre 5 i 30 μm . L’exactitud d’aquest mecanisme és tal que una serp de cascavell cega pot orientar el seu cop a les parts del cos vulnerables d’una presa. [26] Anteriorment es pensava que els òrgans termals evolucionaven principalment com a detectors de preses, però ara es creu que també es poden utilitzar per controlar la temperatura corporal. [27]
Els fosses de radiació tèrmica van experimentar una evolució paral·lela en els cròtals i en alguns boes i pitons, evolucionant també en crotalins i en diverses ocasions en boas i pitons. [28] L’ electrofisiologia de l’estructura és similar entre llinatges, però difereix en l'estructura anatòmica grossa. En un espai més superficial, els crotalinis posseeixen un òrgan termal a cada banda del cap, entre l’ull i la fosa nasal (foseta loreal ). Per la seva part, les boes i els pitons tenen tres o més fosses comparativament més petites que revesteixen el llavi superior i, de vegades, el llavi inferior, dins o entre les escates. Les dels crotalins són els més avançats, amb una membrana sensorial en suspensió en contraposició a una estructura simple estructura de fossa tèrmica. Dins la família de Viperids, l’òrgan termal només es veu a la subfamília Crotalinae : els crotalins. Malgrat la detecció de radiació IR, el mecanisme IR de les fosses no és igual als fotoreceptors. Mentre que els fotoreceptors detecten la llum mitjançant reaccions fotoquímiques, la proteïna que es troba a les fosses facials de les serps és un canal iònic sensible a la temperatura. Aquest canal, permet els senyals d’infrarojos mitjançant un mecanisme que comporta l’escalfament de l’òrgan termal, en lloc d’una reacció química a la llum. [29] Es un mecanisme coherent amb la fina membrana de fossa, que permet que la radiació IR entri a escalfar de forma ràpida i precisa un canal iònic determinat, i desencadeni un impuls nerviós, a més de vascularitzar la membrana de fossa que refreda ràpidament el canal iònic de nou al seu "descans" o "desactivant" la temperatura.
Els vampirs comuns ( Desmodus rotundus ) tenen uns sensors IR especialitzats en en el seu òrgan nasal. [30] Aquests ratpenats són els únics mamífers que s’alimenten exclusivament de sang. El sentit IR permet els premet localitzar animals homeotèrmics com ara bestiar i cavall en un rang d’entre 10 i 15 cm. Aquesta percepció d’infrarojos en vampirs es pot utilitzar per detectar regions de fluxe màxim de sang en els les preses objectives.
Autocomunicació
[modifica]L'autocomunicació és un tipus de comunicació en què l’emissor i el receptor són el mateix individu. L’emissor emet un senyal que està alterat per l’entorn i finalment és rebut pel mateix individu. El senyal alterat proporciona informació que pot indicar aliments, depredadors o altres coespecificitats. Com que l'emissor i el receptor són el mateix animal, la pressió selectiva maximitza l'eficàcia del senyal. Es a dir, el grau en què un senyal emès és identificat correctament per un receptor malgrat la distorsió i el soroll de la propagació. Hi ha dos tipus d’autocomunicacions. El primer és l’ electrorecepció activa que es troba en els peixos elèctrics Gimnotiformes (peixos ganivet) i Mormirids (peixos elefant) i també en el ornitorrinc (Ornithorhynchus anatinus ). El segon tipus d’autocomunicacions és l’ecolocalització, que es troba en ratpenats i balenes dentades .
Funcions
[modifica]Hi ha moltes funcions de comunicació animal. Tot i això, algunes s’han estudiat amb més detall que d’altres. Podem veure:
- Comunicació durant les competicions: La comunicació animal té un paper fonamental en la determinació del guanyador en una competició sobre un recurs. Moltes espècies presenten senyals diferents que indiquen agressió, voluntat d’atacar, o senyals per transmetre la rendició durant les competicions per aliments, territorials o per cercar parella.
- Rituals d'aparellament : Els animals emeten senyals per atraure l’atenció d’una possible parella o per enfortir-ne els lligams. Aquests senyals freqüentment impliquen la visualització de parts o postures del cos. Per exemple, una gasela assumirà posicions característiques per iniciar l'aparellament. Els senyals d’aparellament també poden incloure l’ús de senyals olfactius o sons d'aparellament exclusius d’una espècie. Els animals que formen vincles de parella duradors sovint tenen visualitzacions simètriques que es fan entre si. Exemples cèlebres són la presentació mútua de tiges de canya per part del Cabussó emplomallat que ha estudiat Julian Huxley. També podem veure les exhibicions de triomf mostrades per moltes espècies d'oques i pingüins en els seus llocs niu, i l’espectacular exhibició de festeig que fan les ocells del paradís.
- Propietat / territori : Senyals que s’utilitzen per reclamar o defensar un territori, menjar o la parella.
- Senyals relacionades amb els aliments : Molts animals fan "senyals d'alimentació" per atraure la parella, descendència o altres membres d'un grup social cap a una font d'alimentació. Potser el senyal més elaborat relacionat amb els aliments és la dansa de les abelles de les abelles de la mel estudiades per Karl von Frisch. Un dels exemples coneguts de la captació de la descendència en un niu són els ocells d'espècies altricials (neixen cecs i sords). Els corbs petits assenyalen als corbs adults quan es trobin amb menjar nou o que no s'ha provat. Els macacos Rhesus envien senyals d'aliments per informar a altres membres del grup d'una font d'alimentació per evitar càstigs. Molts insectes socials alliberen feromones per conduir els altres membres del grup a la font d'aliments. Per exemple, les formigues deixen un rastre de feromones a terra que poden ser seguides per altres formigues i dur-les a la font d'alimentació.
- Senyals d’alarma : Les senyals d’alarma comuniquen l’amenaça d’un depredador. Això permet que tots els membres d’un grup social (i de vegades altres espècies) responguin en conseqüència. Això pot incloure córrer per amagar-se, quedar immòbil, o agrupar-se en un per reduir el risc de l’atac. Els senyals d’alarma no sempre són vocalitzacions. Les formigues aixafades alliberaran una feromona d’alarma per atraure noves formigues i enviar-les un estat d’atac.
- Meta-comunicació : Son senyals que modificaran el significat dels senyals posteriors. Un exemple és la “simulació” en els gossos que indiquen un senyal agressiu posterior forma part d’una baralla de joc en lloc d’un episodi greu d'agresivitat.
Interpretació del comportament dels animals
[modifica]Com s'ha descrit anteriorment, molts gestos, postures i sons d'animals transmeten un significat als animals propers. Aquests senyals són sovint més fàcils de descriure que d’interpretar. És temptador, sobretot amb animals i simis domesticats, antropomorfitzar. Es a dir, interpretar accions dels animals en termes humans. Però això pot resultar força enganyós. Per exemple, el "somriure" d'un simi sovint és un signe d'agressió. A més, el mateix gest pot tenir significats diferents segons el context en què es produeixi. Per exemple, la postura de la cua d’un gos domèstic es pot utilitzar de diferents maneres per transmetre molts significats tal com s’il·lustra a L’Expressió i les emocions en l’home i els animals de Charles Darwin publicada el 1872. Algunes de les il·lustracions de Darwin les reproduim aquí.
- Examples of tail position indicating different emotions in dogs
-
 "Petit gos veient un gat sobre una taula"
"Petit gos veient un gat sobre una taula" -
 "Gos que s'apropa a un altre gos amb intencions hostils"
"Gos que s'apropa a un altre gos amb intencions hostils" -
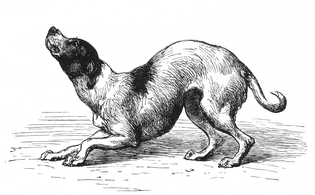 "Gos amb un marc mental humil i afectuós"
"Gos amb un marc mental humil i afectuós" -
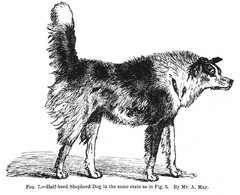 "Can pastor mig criat"
"Can pastor mig criat" -
 "Gos acaricia el seu amo"
"Gos acaricia el seu amo"





Comunicació entre espècies
[modifica]Molta comunicació animal és dona entre membres de diferents espècies. Pel que fa a aquest tipus de comunicació, te una especial interès aquella que succeeix entre el depredador i la seva presa.
Presa a depredador
[modifica]Si un animal de presa es mou, fa un soroll, vibra, o emet un olor de manera que un depredador pugui detectar-lo, aquest fet es coherent amb la definició de "comunicació" indicada anteriorment. Aquest tipus de comunicació es coneix com a interrupció d'una escolta amagada, quan resulta quan un depredador intercepta un missatge destinat a aspectes espècies.
No obstant això, hi ha algunes accions d’espècies que son presa que estan clarament dirigides a depredadors reals o potencials. Un bon exemple és aposematisme: espècies com les vespes són capaces de perjudicar possibles depredadors sovint amb colors vius. Aquesta reacció modifica el comportament del depredador, que instintivament o per l’experiència acumulada, evitarà atacar un animal d’aquest tipus. Algunes formes de mimetisme entren en la mateixa categoria: per exemple, la família dels sirfids s'acoloreixen de la mateixa manera que les vespes i, encara que no són capaços de picar, la forta precaució vers les vespes per part dels seus depredadors proporciona els hi proporciona una protecció. També hi ha canvis de conducta que actuen d’una manera similar en els avisos per coloració. Per exemple, canins com els llops i els coiots poden adoptar una postura agressiva, com ara gronxar-se amb les dents bloquejades, per indicar que lluitaran si es necessari. Les serps cascavells utilitzen el seu conegut cascavell per avisar els possibles depredadors de la seva picada verinosa. De vegades, es pot combinar amb un canvi de comportament i una coloració d’alerta, com en certes espècies d’amfibis que tenen la major part del seu cos que es mimetitza amb el seu entorn, excepte el ventre que te colors vius. Quan s’enfronten a una amenaça potencial, mostren el ventre, cosa que indica que d’alguna manera són verinosos.
Un altre exemple de comunicació de presa a predador és el senyal de persecució-dissuasió. Els senyals de persecució-dissuasió es produeixen quan les preses indiquen a un depredador que la caça no seria rendible perquè l'espècie que emet el senyat està preparada per escapar. Els senyals de persecució-dissuasió proporcionen un avantatge tant per l'emissor com per al receptor: impedeixen que l’emissor perdi temps i energia fugint i impedeixin que el receptor inverteixi en una tasca costosa que és poc probable que tingui èxit en la captura. Aquests senyals poden anunciar la capacitat d’escapament de les preses i reflectir l’estat fenotípic (qualitat del senyal) o poden anunciar que la presa ha detectat el depredador (senyal de percepció). S’han reportat senyals de persecució-dissuasió en una gran varietat de tàxons, inclosos peixos (Godin i Davis, 1995), llangardaixos (Cooper et al., 2004), ungulats (Caro, 1995), conills (Holley 1993), primats (Zuberbuhler et al. 1997), rosegadors (Shelley i Blumstein 2005, Clark, 2005), i ocells (Alvarez, 1993, Murphy, 2006, 2007). Un exemple familiar de senyal qualitatiu de persecució-dissuasió és el salt de rebot, una combinació repetida de salts verticals amb les quatre potes rígides mentre van corrent en alguns antílops com la gasela de Thomson en presència d’un depredador. S’han proposat, com a mínim, 11 hipòtesis per aquest comportament. La teoria majorment acceptada en l'actualitat és que alerta als depredadors que han perdut la capacitat de sorprendre. Depredadors com els guepards, que confien en atacs sorpresa, demostrat pel fet que les persecucions rarament tenen èxit quan els antílops mostren aquest comportament. Els depredadors no malgasten energia en una persecució que probablement no tindrà èxit (comportament òptim de cerca d'aliment). Els senyals qualitatius es pot comunicar amb altres modes que no siguin visuals. La rata cangur produeix diversos patrons de percussió de peus en diversos contextos, un dels quals es quan es troba amb una serp. La percussió amb els peus pot alertar als descendents que estiguin propers, però el més probable és que transmeti vibracions a través del sòl que la rata està massa alerta per tenir un atac amb èxit, evitant així la persecució depredadora de la serp. [31]
Predador a presa
[modifica]
Normalment, els depredadors intenten reduir la comunicació a les preses, ja que generalment redueixen l'efectivitat de la seva caça. Tot i això, algunes formes de comunicació de predadors a les preses tenen lloc de maneres que canvien el comportament de la presa i faciliten la seva captura. Es a dir, el depredador enganya a la seva presa de forma intencionada per obtenir-ne la seva captura. Un dels casos mes coneguts és el peixos lofiformes, que actuen com un depredador emboscat que espera a que arribi la seva. Té un creixement carnós bioluminescent que sobresurt del seu front i que desemboca a les seves mandíbules. Els peixos més petits intenten agafar l'esquer, situant-se en una posició millor per a que el predador els agafi. Un altre exemple de comunicació enganyosa s’observa en el gènere d’aranyes saltadores ( Myrmarachne ). Aquestes aranyes es coneixen habitualment com a "aranyes anti-mimecomorfisme " a causa de la manera de mimetitzar les potes anteriors a l'aire per simular les antenes .
Comunicació Home-animal
[modifica]La comunicació entre espècies, en el cas d'humans i animals, permet entendre com els humans interpreten el comportament dels animals o els hi donen ordres . Una aspecte rellevant de les comunicacions humans-animals pot ser clau pel al benestar dels animals que són cuidats o ensinistrats pels humans. Per exemple, cal reconèixer conductes que indiquen dolor. De fet, la supervivència tant de l'animal com del seu cuidador humà pot estar en joc si, per exemple, un humà no reconeix un senyal per a un atac imminent. També és important tenir en compte que les espècies d’animals no humans poden interpretar els senyals dels humans d’una altra manera que els propis humans. Per exemple, una ordre de comandament fa referència mes a una ubicació que a un objecte en els gossos. [32]
Des de finals dels anys 90, l'investigador Sean Senechal, desenvolupa, estudia i utilitza el llenguatge visible i expressiu après en gossos i cavalls. A l'ensenyar a aquests animals un llenguatge de signes, similar al llenguatge de signes americà, s'ha trobat que els animals utilitzen els nous signes pel seu compte per obtenir el que necessiten. [33] Els experiments recents sobre llenguatge animal són potser l’intent més sofisticat que encara s’ha d’establir la comunicació humans-animals, tot i que la relació amb la comunicació natural dels animals és incerta.
Altres aspectes
[modifica]Evolució
[modifica]La importància de la comunicació esta vinculada a la morfologia, el comportament i la fisiologia que han elaborat alguns animals per facilitar el seu desenvolupament. Algunes de les estructures més marcades del regne animal, les trobem en la cua del paó, les banyes d'un cérvol, el pit del llangardaix de gorguera, o la modesta taca vermella del gavià argentat europeu. S'han desenvolupat comportaments molt elaborats per a la comunicació com el ball de grues, els canvis de color de les sèpies, o la recollida i organització de materials per part dels ocells arquers . Una altra evidència de la importància de la comunicació en els animals és la priorització de les funcions fisiològiques a aquesta funció, En aquest sentit, sembla que la vocalització dels ocells tenen estructures cerebrals totalment dedicades a la seva producció. Totes aquestes adaptacions requereixen una explicació evolutiva.
Hi ha dos aspectes a considerar per aquesta explicació:
- identificar com un animal al que li mancava un tret característic del comportament rellevant pogués adquirir-lo;
- identificar la pressió selectiva que el comportament adaptatiu en animals pugui desenvolupar estructures que facilitin la comunicació, emeten senyals i responent a aquestes.
Konrad Lorenz i altres primers etòlegs van fer contribucions importants al primer d'aquests problemes. En comparar espècies relacionades en un grup, van demostrar que els moviments i les parts del cos que, en les formes primitives, no tenien cap funció comunicativa, podrien ser "capturats" en un context on la comunicació fos funcional per a un o els dos membres i pogués evolucionar cap a una forma més elaborada i especialitzada. Per exemple, Desmond Morris va mostrar en un estudi sobre els fringil·lids que els canvis en el seu bec tenia repercussions en diverses espècies, servint com una funció de empolainament, tot i que en algunes espècies aquesta havia estat elaborada en un senyal de festeig .
El segon problema ha estat més controvertit. Els primers etòlegs van suposar que la comunicació es produïa per al bé de l’espècie en general. Això requeriria un procés de selecció de grups que es creu matemàticament impossible en l’ evolució de la reproducció sexual dels animals. L’altruisme cap a un grup no relacionat no s’accepta àmpliament a la comunitat científica. Es pot veure com un altruisme recíproc, esperant el mateix comportament dels altres, sobre el benefici de viure en grup. Els sociobiòlegs van argumentar que podrien sorgir comportaments que beneficiaven un grup sencer d’animals com a resultat de pressions de selecció que actuaven únicament sobre l’individu. La Teoria de l'evolució centrada en el gen proposa que els comportaments que permeten a un gen establir-se en una població més àmplia dins d'una població s'haurien seleccionat positivament, fins i tot si el seu efecte sobre els individus o l'espècie en conjunt resultés perjudicial. [34]

En el cas de la comunicació, una discussió important entre John Krebs i Richard Dawkins va establir hipòtesis sobre l'evolució de les comunicacions tan aparentment altruistes o mutualistes com les senyals d'alarma i de festeig, apareixent sota la selecció individual. Això va suposar que la comunicació no sempre seria "honesta" (de fet, hi ha alguns exemples evidents on no ho és, com passa en el mimetisme ). La possibilitat d'una comunicació deshonesta evolutivament estable ha estat objecte de molta controvèrsia. Amb Amotz Zahavi, en particular, defensant que no pot existir a llarg termini. Els sociobiòlegs també s’han preocupat per l’evolució d’estructures de senyalització aparentment excessives com la cua del paó. Es pensa que aquestes només poden sorgir com a resultat de la selecció sexual. Per aquest motiu, es pot generar un procés de retroalimentació positiva, que condueix a la ràpida exageració d'una característica que confereix un avantatge en una situació de selecció de parella competitiva.
Una teoria per explicar l'evolució de trets com la cua d'un paó és la "selecció desbocada". Això requereix dos elements: un factor que existeix, com la cua brillant, i un biaix preexistent en la femella per seleccionar-ne el tret. Les femelles prefereixen les cues més elaborades i, per tant, aquests mascles són capaços d’aparellar-se amb èxit. Aprofitant la psicologia de la femella, es produeix un bucle de retroalimentació positiva i la cua es fa més gran i brillant. Finalment, l'evolució s'anivellarà perquè els costos de supervivència del mascle no permeten que s'elabori aquest element. Existeixen dues teories per explicar la selecció desbocada. El primer és la hipòtesi d'uns bons gens. Aquesta teoria estableix que una visualització elaborada és un senyal honest, de manera que es pot escollir el la millor parella. El segon és la hipòtesi del handicap. Aquesta explica que la cua del paó sigui un handicap, que requereix energia per mantenir-la i la fa més visible per als depredadors. Per tant, el senyal és costós de mantenir i continua sent un indicador honest per part de l'emissor del senyal. Un altre supòsit és que el senyal és més costós per produir mascles de baixa qualitat que per produir mascles de més qualitat. Això es deu simplement a què individus de major qualitat del sexe masculí tenen més reserves d’energia disponibles per assignar a una senyalització costosa. [1]
Aspectes cognitius
[modifica]Els etòlegs i sociobiòlegs han analitzat les característiques de la comunicació animal en termes de respostes més o menys automàtiques als estímuls, sense plantejar-se si els animals afectats comprenen el significat dels senyals que emeten i reben. Aquesta és una qüestió clau de la cognició animal . Hi ha alguns sistemes de senyalització que semblen exigir una comprensió més avançada. Un exemple molt discutit és l’ús dels senyals d’alarma per part de les mones vervet . Robert Seyfarth i Dorothy Cheney van demostrar que aquests animals emeten diferents senyals d’alarma en presència de diferents depredadors (lleopards, àguiles i serps) i que les mones que escolten els senyals responen de forma adequada. Però aquesta habilitat es desenvolupa amb el pas del temps i, a més, cal tenir en compte l’experiència de la persona que emet el senyal. La metacomunicació, comentada anteriorment, també necessita un procés cognitiu més sofisticat.
Investigacions recents sobre els dofins nas de botella[35] demostren que poden reconèixer la identitat de la informació dels dels xiulets, fins i tot quan son contraris a les característiques del senyal. Aquests dofins son els únics animals diferents als humans que demostren que transmeten informació d'identitat independent de la veu o de la ubicació de l'emissor. L’article conclou que:
| « | El fet que la forma del xiulet del dofí porti informació d’identitat independent de les funcions de veu presenta la possibilitat d’utilitzar aquests xiulets com a senyals referencials, ja sigui dirigint-se a individus o referint-se a ells. De forma similar a l’ús de noms en humans. Tenint en compte les habilitats cognitives dels dofins nas de botella, els seus aprenentatges vocals, les seves habilitats bucals, i la seva estructura social de fisió-fusió, es tracta d'un factor del qual encara calen mes investigacions. | » |
| — V. M. Janik, et al. [35] | ||
Comportament humà
[modifica]Un altre tema controvertit és fins a quin punt els comportaments humans s’assemblen a la comunicació animal, o si tota aquesta comunicació ha desaparegut com a resultat de la nostra capacitat lingüística. Algunes de les nostres característiques corporals: celles, barbes i bigotis, veus masculines adultes profundes, potser pits femenins, s’assemblen molt a les adaptacions per produir senyals. Etòlegs com Irenäus Eibl-Eibesfeldt han defensat que els gestos facials com somriure, disgust, i l'arqueig de celles en la salutació són senyals comunicatives humanes universals que es poden relacionar amb els senyals corresponents en altres primats. Tenint en compte la forma en què ha aparegut el llenguatge parlat recentment, és molt probable que el llenguatge corporal humà inclogui respostes més o menys involuntàries que tinguin un origen similar a la comunicació que tenim.
Els humans també busquen imitar els senyals comunicatius dels animals per interactuar amb ells. Per exemple, els gats tenen una resposta de fidelitat lleu al tancar lentament els ulls. Els humans sovint imiten aquest senyal cap a un gat de animal de companyia per establir una relació de tolerància. Acariciar, acaronar i fregar els animals de companyia són accions que probablement funcionen mitjançant els seus patrons naturals de comunicació interespecífica.
Els gossos han demostrat la capacitat d’entendre la comunicació humana. En les tasques d’elecció d’objectes, els gossos utilitzen gestos comunicatius humans, com apuntar i dirigir la mirada per localitzar aliments i joguines ocults. [36] També s’ha demostrat que els gossos presenten una mirada esquerra quan miren les cares humanes, cosa que indica que són capaços de llegir les emocions humanes. [37] Els gossos no fan ús de la direcció de la mirada ni mostren la direcció esquerra amb la mirada amb altres gossos.
Un nou enfocament al segle XXI en el camp de la comunicació animal utilitza anàlisis conductuals aplicat, en un entrenament específic funcional en la comunicació. Aquesta forma de formació prèviament s’ha utilitzat a les escoles i clíniques amb humans amb necessitats especials, com els nens amb autisme, per ajudar-los a desenvolupar el llenguatge. Sean Senechal, al Centre AnimalSign, ha estat utilitzant un enfocament similar a l’entrenament funcional de comunicació amb animals domèstics, com ara gossos des del 2004 i cavalls des del 2000, amb resultats positius tant per animar animals com per les persones. L’ensinistrament de comunicació funcional per a animals, Senechal l'anomena “AnimalSign Language”. Això inclou l’ensenyament de la comunicació mitjançant gestos (com el llenguatge de signes americà simplificat), el PECS (sistema de comunicació per intercanvi d’imatges), el punteig i la vocalització. El procés per als animals inclou tècniques simplificades i modificades. [ <span title="This claim needs references to reliable sources. (August 2018)">cita requerida</span> ]
Lingüística
[modifica]| Do animals have language? - Michele Bishop, TED Ed, 4:54, September 10, 2015[38] | |
Per a la lingüística, l'interès dels sistemes de comunicació animal rau en les seves similituds i diferències amb el llenguatge humà:
- Les llengües humanes es caracteritzen per tenir una doble articulació (en la caracterització del lingüista francès André Martinet ). Vol dir que es poden desglossar expressions lingüístiques complexes en elements significatius (com ara morfemes i paraules ), que a la vegada estan compostos per elements fonètics més petits que afecten el significat, anomenats fonemes . Els senyals animals, però, no presenten aquesta doble estructura.
- En general, les expressions dels animals són respostes a estímuls externs i no fan referència a qüestions eliminades en el temps i l'espai. Les qüestions rellevants a distància, com ara fonts d'alimentació llunyanes, solen indicar-se a altres individus mitjançant el llenguatge corporal. Per exemple, l'activitat del llop abans de caçar, o la informació que es transmet en un llenguatge de dansa d'abelles . Per tant, no se sap fins a quin punt les intervencions són respostes automàtiques i fins a quin punt hi ha una intenció intencionada.
- En contrast amb el llenguatge humà, els sistemes de comunicació animal no solen expressar generalitzacions conceptuals. (Els cetacis i alguns primats poden ser excepcions notables).
- Els llenguatges humans combinen elements per produir nous missatges (propietat coneguda com a creativitat ). Un dels factors és que el creixement del llenguatge humà es basa en idees conceptuals i estructures hipotètiques, ambdues capacitats molt més grans en els humans que en els animals. Això sembla molt menys freqüent en els sistemes de comunicació animal, tot i que la investigació actual sobre la cultura animal continua sent un procés en progressió amb molts nous descobriments.
Errors en la comunicació
[modifica]Hi ha possibilitat d’error en la comunicació entre animals quan es donen determinades circumstàncies. [39] Aquestes circumstàncies podrien incloure la distància entre els dos subjectes que es comuniquen, així com la complexitat del senyal que s’està comunicant al receptor de la situació. Pot ser que no sempre sigui clar pel receptor d'on prové la ubicació de la comunicació, ja que l'emissor de vegades pot enganyar-lo i crear més errors. [ <span title="This claim needs references to reliable sources. (October 2018)">cita requerida</span> ]
Vegeu també
[modifica]- Consciència dels animals
- Antrozoologia (estudis relació home-animal)
- Biocomunicacions
- Biosemiòtics
- Llenguatge corporal (cinèsica)
- Estimat efecte enemic i Efecte veí Nasty
- L’engany en animals
- Degeneració (biologia)
- Emoció en animals
- Relacions socials
- Comunicació entre humans i animals
- International Society for Biosemiotic Studies
- Origen de la llengua
- Origen del discurs
- Joc de Sir Philip Sidney
- Animal parlant
- Zoomusicologia
- Zoosemiòtics
Referències
[modifica]- ↑ 1,0 1,1 Maynard-Smith and Harper, 2003
- ↑ de Waal
- ↑ Langford, D.J. et al., (2010). Coding of facial expressions of pain in the laboratory mouse. Nature Methods, May 9th. pp. 1–3. doi:10.1038/nmeth.1455
- ↑ Bräuer, Juliane; Call, Josep; Tomasello, Michael (en anglès) Journal of Comparative Psychology, 119, 2, 2005, pàg. 145–154. DOI: 10.1037/0735-7036.119.2.145. ISSN: 1939-2087. PMID: 15982158.
- ↑ Téglás, Ernő; Gergely, Anna; Kupán, Krisztina; Miklósi, Ádám; Topál, József Current Biology, 22, 3, 2012, pàg. 209–212. DOI: 10.1016/j.cub.2011.12.018. ISSN: 0960-9822. PMID: 22226744.
- ↑ Schloegl, Christian; Kotrschal, Kurt; Bugnyar, Thomas Animal Behaviour, 74, 4, 2007, pàg. 769–778. DOI: 10.1016/j.anbehav.2006.08.017. ISSN: 0003-3472.
- ↑ Range, F.; Virányi, Z. PLoS ONE, 6, 2, 2011, pàg. e16888. Bibcode: 2011PLoSO...616888R. DOI: 10.1371/journal.pone.0016888. PMC: 3044139. PMID: 21373192.
- ↑ Cloney, R.A.; Florey, E. Z. Zellforsch Mikrosk. Anat., 89, 2, 1968, pàg. 250–280. DOI: 10.1007/bf00347297. PMID: 5700268.
- ↑ Hanlon, R.T.. Cephalopod Behaviour. Cambridge University Press, 1996, p. 121. ISBN 978-0-521-64583-6.
- ↑ Motluk, Alison New Scientist, 19, 7, 2001.
- ↑ Ehrlich, Paul R.. «"Bird Voices" and "Vocal Development" from Birds of Stanford essays». [Consulta: 9 setembre 2008].
- ↑ Slabbekoorn, Hans, Smith, Thomas B. "Bird song, ecology and speciation." Philosophical Transactions: Biology Sciences 357.1420 (2002). 493–503.
- ↑ Zuberbühler, Klause. "Predator-specific alarm calls in Campbell's monkeys, Cercopithecus campbelli." Behavioral Ecology and Sociobiology 50.5 (2001). 414–442
- ↑ Boughman, Janette W. "Vocal learning by greater spear-nosed bats." Proceedings: Biological Sciences 265.1392 (1998). 227–233
- ↑ Krulwich, Robert. «New Language Discovered: Prairiedogese». [Consulta: 20 maig 2015].
- ↑ DeMello, Margo. «Yips, barks and chirps: the language of prairie dogs», 2007. [Consulta: 20 maig 2015].
- ↑ «Prairie dogs' language decoded by scientists». CBC News, 21-06-2013. [Consulta: 20 maig 2015].
- ↑ Carey, Bjorn. Whales Found to Speak in Dialects. Live Science. 3 Jan. 2006.
- ↑ 19,0 19,1 Bradbury, J.W., and S.L. Vehrencamp. Principles of Animal Communication. Sunderland, MA: Sinauer Associates Inc., 2011. Print.
- ↑ Brown, G.E.; Chivers, D.P.; Smith, R.J.F. Behavioral Ecology and Sociobiology, 36, 2, 1995, pàg. 105–110. DOI: 10.1007/s002650050130.
- ↑ Gosling, L. Morris, and S. Craig Roberts. "Scent-marking by male mammals: cheat-proof signals to competitors and mates." Advances in the Study of Behavior 30 (2001): 169–217.
- ↑ du P. Bothma, J.; Richet, E.A.N. le Journal of Arid Environments, 29, 4, 1995, pàg. 511–517. Bibcode: 1995JArEn..29..511D. DOI: 10.1016/s0140-1963(95)80023-9.
- ↑ «Electrocommunication». Davidson College. Arxivat de l'original el 2012-05-29. [Consulta: 3 març 2011].
- ↑ Bradbury, J.W., and S.L. Vehrencamp. Principles of Animal Communication. Sunderland, MA: Sinauer Associates Inc., 2011.
- ↑ Narins, Peter M. "Seismic Communication in Anuran Amphibians." BioScience40.4 (1990): 268. Print.
- ↑ (Kardong & Mackessy 1991)
- ↑ (Krochmal et al. 2004)
- ↑ (Pough et al. 1992)
- ↑ (Gracheva et al. 2010)
- ↑ Kürten, L., Schmidt, U. and Schäfer, K. Naturwissenschaften, 71, 6, 1984, pàg. 327–328. Bibcode: 1984NW.....71..327K. DOI: 10.1007/BF00396621.
- ↑ «Web of Life:Vibrational communication in animals». [Consulta: 8 desembre 2012].
- ↑ Tauzin, Tibor; Csík, Andor; Kis, Anna; Topál, József (en anglès) Journal of Comparative Psychology, 129, 4, 2015, pàg. 334–338. DOI: 10.1037/a0039462. ISSN: 1939-2087.
- ↑ Sean Senechal: Dogs can sign, too. A breakthrough method of teaching your dog to communicate to you, 2009, Random House/Crown/TenSpeed Press
- ↑ discussed at length by Richard Dawkins under the subject of his book The Selfish Gene
- ↑ 35,0 35,1 V. M. Janik, L. S. Sayigh, and R. S. Wells: "Signature whistle shape conveys identity information to bottlenose dolphins", Proceedings of the National Academy of Sciences, vol. 103 no 21, May 23, 2006
- ↑ Hare, B.; Call, J.; Tomasello, M. Evolution of Communication, 2, 1998, pàg. 137–159. DOI: 10.1075/eoc.2.1.06har.
- ↑ K. Guo, K. Meints, C. Hall, S. Hall & D. Mills: "Left gaze bias in humans, rhesus monkeys and rhesus domestic dogs." "Animal Cognition", vol. 12, 2009
- ↑ «Do animals have language? - Michele Bishop». TED Ed, 10-09-2015. [Consulta: 11 setembre 2015].
- ↑ Real, Leslie A. Behavioral Mechanisms in Evolutionary Ecology. University of Chicago Press, 1994-12-15. ISBN 9780226705958.
Enllaços externs
[modifica]- Animal Communicator – Documental
- Zoosemiotics: animal communication on the web
- The Animal Communication Project
- International Bioacoustics Council recerca sobre llenguatge animal.
- Animal Sounds different sons animals per escoltar i descarregar .
- The British Library Sound Archive incorpora sobre 150,000 gravacions de sons d'animals i ambients naturals de tot el mon.
[[Categoria:Semiòtica]] [[Categoria:Pàgines amb traduccions sense revisar]]