
Eocè
| Període | Època | Estatge | Edat (Ma) | |
|---|---|---|---|---|
| Neogen | Miocè | Aquitanià | (més recent) | |
| Paleogen | Oligocè | Catià | 27,82 | |
| Rupelià | 33,9 | |||
| Eocè | Priabonià | 37,2 | ||
| Bartonià | 40,4 | |||
| Lutecià | 48,6 | |||
| Ipresià | 55,8 | |||
| Paleocè | Thanetià | 58,7 | ||
| Selandià | 61,6 | |||
| Danià | 66,0 | |||
| Cretaci | Superior | Maastrichtià | (més antic) | |
| Comissió Internacional d'Estratigrafia (2023) | ||||
L'Eocè[nota 1] o Eocé[nota 2] és l'època geològica que començà fa 56,0 milions d'anys i s'acabà fa 33,9 milions d'anys. Es tracta de la segona època del període Paleogen i de l'era del Cenozoic, que encara dura. El seu nom deriva de les paraules gregues antigues ἠώς (eós), que significa ‘alba’, i καινός (kenós), que significa ‘recent’, i en el seu conjunt es pot traduir per ‘alba dels temps recents’.[1]
El seu inici i final queden marcats pel màxim tèrmic del Paleocè-Eocè i per la Grande Coupure, respectivament. Com en altres períodes geològics, els estrats que delimiten aquest període estan ben identificats, però no han estat datats amb màxima precisió.
Subdivisions
[modifica]L'Eocè se sol subdividir en Eocè inferior (Ipresià), Eocè mitjà (Lutecià+Bartonià) i Eocè superior (Priabonià). De manera menys habitual, també es pot subdividir únicament en inferior i superior. En aquest cas, l'estatge Lutecià passa a formar part de l'Eocè inferior i l'estatge Bartonià passa a formar part de l'Eocè superior.
Els quatre estatges faunístics de l'Eocè són:
- Ipresià: començà fa 55,8 ± 0,2 milions d'anys amb l'inici del màxim tèrmic del Paleocè-Eocè, un període de ràpid i intens escalfament global que provocà l'extinció de nombrosos foraminífers bentònics, però que coincidí amb un episodi de gran evolució i radiació dels mamífers. Estratigràficament, el seu inici queda marcat per la base d'una excursió negativa d'isòtops de carboni. El seu final queda marcat per foraminífers planctònics i l'aparició inferior del gènere Hantkenina. S'acabà fa 48,6 ± 0,2 milions d'anys i el seu nom, encunyat el 1850 pel geòleg belga André Hubert Dumont, fa referència a la localitat d'Ypres (Bèlgica).[2]
- Lutecià: començà fa 48,6 ± 0,2 milions d'anys. És un període molt ric en invertebrats marins (mol·luscs, coralls o eriçons de mar) i que es caracteritza per l'abundància de mars epicontinentals soms sotmesos a les influències continentals. Estratigràficament, el seu inici queda marcat per foraminífers planctònics i l'aparició inferior del gènere Hantkenina. El seu final queda marcat per la quasiextinció del nanofòssil calcari Reticulofenestra reticulata. S'acabà fa 40,4 ± 0,2 milions d'anys i el seu nom, encunyat el 1883 pel geòleg francès Albert de Lapparent, fa referència a l'antic nom romà de París, Lutècia.[3]
- Bartonià: començà fa 40,4 ± 0,2 milions d'anys. El seu nom li fou donat per Karl Mayer-Eymar, que en definí els límits l'any 1857 a partir de sediments argilosos del sud d'Anglaterra rics en fòssils. Estratigràficament, el seu inici queda marcat per la quasiextinció del nanofòssil calcari Reticulofenestra reticulata. El seu final queda marcat per una de les aparicions més inferiors del nanofòssil calcari Chiasmolithus oamaruensis. S'acabà fa 37,2 ± 0,2 milions d'anys i el seu nom, encunyat el 1857 pel geòleg alemany Karl Mayer-Eymar, fa referència a la localitat de Barton on Sea (New Milton) (Anglaterra).[4]
- Priabonià: començà fa 37,2 ± 0,1 milions d'anys. És l'últim estatge faunístic de l'Eocè, i la Grande Coupure, un episodi d'extincions, de canvis faunístics i d'abundant especiació, tingué lloc a finals del període. Estratigràficament, el seu inici queda marcat per una de les aparicions més inferiors del nanofòssil calcari Chiasmolithus oamaruensis. El seu final queda marcat per foraminífers planctònics i l'extinció del gènere Hantkenina. S'acabà fa 33,9 ± 0,1 milions d'anys i el seu nom, encunyat el 1893 pels geòlegs francesos Ernest Munier-Chalmas i Albert de Lapparent, fa referència a la localitat de Priabona (Itàlia).[5]
Paleoclima
[modifica]
El clima global de l'Eocè fou probablement el més homogeni del Cenozoic; el gradient tèrmic de l'equador als pols era la meitat de l'actual, i els corrents oceànics profunds eren excepcionalment càlids. Les regions polars eren molt més càlides que avui en dia, potser com el nord-oest dels Estats Units actual. Els boscos temperats arribaven fins als mateixos pols, mentre que els climes tropicals plujosos arribaven fins a 45° de latitud nord. La diferència era més gran a les latituds temperades; tanmateix, el clima dels tròpics probablement era similar al de l'actualitat.[6] Com que el continent australià i l'Antàrtida encara estaven units en una sola massa terrestre, els corrents oceànics freds i càlids es mesclaven, mantenint una temperatura oceànica homogènia.[7]
Des del principi d'aquest període, la temperatura del planeta augmentà en un dels escalfaments globals més ràpids (en un context geològic) i extrems registrats a la història geològica, anomenat màxim tèrmic del Paleocè-Eocè. Fou un episodi d'escalfament ràpid i intens (de fins a 7 °C a latituds altes) que durà menys de cent mil anys.[8] El màxim tèrmic provocà una gran extinció que serveix per distingir la fauna de l'Eocè de la del Paleocè.
És possible que aquest intens escalfament es degués a l'alliberament de clatrats de metà enterrats al fons marí. Es creu que els sediments en què es trobaven emmagatzemats aquests clatrats foren pertorbats a mesura que els oceans s'escalfaven, i el metà que contenien provocà l'augment de les temperatures globals. S'estima que s'alliberaren unes dues mil gigatones de metà, un gas amb un efecte hivernacle més de deu vegades superior al del diòxid de carboni.[9]

L'augment de les temperatures arreu del món no en fou l'única conseqüència, sinó que el clima global esdevingué més humit. Gran part d'aquesta humitat era conduïda als pols.[10] La gran quantitat de pluja sobre l'oceà Àrtic, juntament amb la configuració dels continents (que l'aïllava de la resta d'oceans), en reduí dràsticament la salinitat.[10] L'aigua dolça acumulada a la zona àrtica preparà el terreny per un gran canvi climàtic de signe totalment oposat i que marcaria el final de l'Eocè.
El clima romangué càlid durant la resta de l'Eocè, tot i que un refredament global que finalment portaria a les glaciacions del Plistocè començà a mitjans de l'època a causa de dos factors: primer l'esdeveniment d'Azolla i després l'aïllament de l'Antàrtida.
L'esdeveniment d'Azolla tingué lloc fa uns 49 milions d'anys, quan creixeren grans quantitats de la falguera d'aigua dolça Azolla a l'oceà Àrtic. A mesura que s'enfonsaven al fons marí, les plantes quedaven incorporades als sediments, on no es descomponien a causa del baix nivell d'oxigen de les capes d'aigua profundes. La reducció resultant de la quantitat de carboni a l'atmosfera terrestre contribuí a transformar el planeta d'una «Terra hivernacle», prou càlida perquè prosperessin tortugues i palmeres als pols, a l'estat actual de «Terra glaçada».[11][12][13]
Amb la seva separació del continent australià fa uns 45 milions d'anys, l'Antàrtida quedà privada del flux d'aigües equatorials que fins aleshores en suavitzaven el clima. Privada d'aquestes aigües càlides, l'Antàrtida es refredà i l'oceà Antàrtic inicià la seva glaciació, creant un flux d'aigua freda (vegeu Corrent circumpolar antàrtic) i banquises que reforçaren l'efecte refredant.
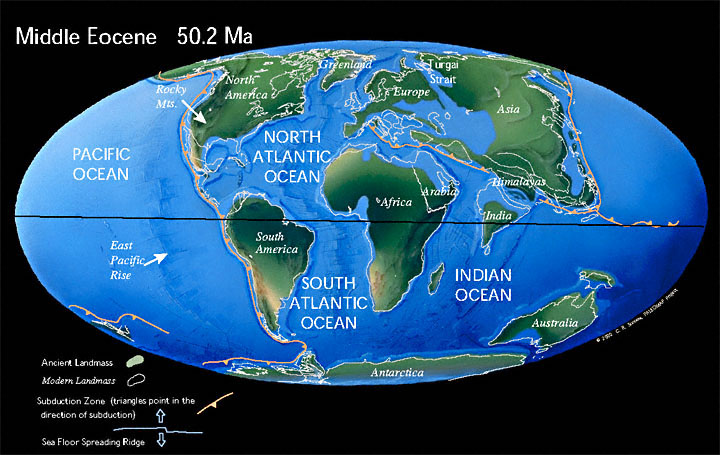
Paleogeografia
[modifica]La tercera i última gran fase de la fragmentació del supercontinent Pangea tingué lloc a principis del Cenozoic, entre el Paleocè i l'Oligocè. Laurèntia (Nord-amèrica) continuà separant-se d'Euràsia i eixamplant el jove oceà Atlàntic, tot i que es creu que encara hi havia una connexió entre ambdues masses terrestres.[14] Mentre l'Atlàntic creixia, l'ancestral oceà de Tetis es continuà tancant a causa de l'aproximació dels continents africà i eurasiàtic.
A principis de l'Eocè, el continent australià i l'Antàrtida encara romanien units, però durant el Lutecià, Austràlia començà a separar-se ràpidament de l'Antàrtida i a derivar cap al nord, tal com ho havien fet el subcontinent indi i Nova Zelanda desenes de milions d'anys abans al període Cretaci. L'aïllament del continent antàrtic tindria conseqüències dràstiques sobre el clima global.
Orogènesi
[modifica]El Terciari fou un període d'intensa activitat orogènica. Durant l'anomenada orogènesi alpina es formaren les muntanyes del sistema de Tetis, una serralada que s'estén per la part meridional d'Euràsia i que inclou els Alps, els Carpats, les muntanyes de l'Àsia Menor i l'Iran, l'Hindu Kush, l'Himàlaia i les muntanyes del sud-est asiàtic.[15] Aquesta orogènesi causà un intens metamorfisme de les roques existents i l'ensorrament d'estrats rocosos.

El subcontinent indi, que s'havia separat de Gondwana al Cretaci superior i s'havia mogut cap al nord a una velocitat de 16 cm/any des d'aleshores, col·lidí amb Euràsia a principis de l'Eocè. La col·lisió entre aquestes dues masses terrestres alçà la serralada més alta del món, l'Himàlaia.[16] Aquest procés d'orogènesi encara dura avui en dia, i fa que l'Himàlaia sigui uns cinc mil·límetres més alt cada any.[17]
D'altra banda, l'orogènesi cimmeriana, un procés orogènic que havia començat al Juràssic, continuà creant algunes de les serralades que actualment es troben al centre del continent asiàtic. L'Eocè fou l'escenari de la fase final d'aquesta orogènesi.
Euràsia no fou l'únic continent amb activitat orogènica. La configuració geològica de nombroses muntanyes d'Amèrica del Nord daten de principis del Cenozoic, com per exemple, les Black Hills de Dakota del Sud i Wyoming, o els Apalatxes de la costa est.
Fauna
[modifica]Aus
[modifica]
Per primera i única vegada en la història de la Terra, els ocells dominaven el món. Aus depredadores gegants com Gastornis (antigament coneguda com a Diatryma) s'alimentaven de mamífers com Propaleotherium a Europa i a Nord-amèrica,[18] mentre que els forusràcids (coneguts com a aus terrorífiques) es convertien en els depredadors alfa d'Amèrica del Sud.
Els pingüins, que havien aparegut durant el Paleocè, arribaren a Amèrica del Sud a l'Eocè mitjà, i al Bartonià ja havien començat a estendre's per aigües de l'Atlàntic. Alguns gèneres i espècies primitius de pingüins són Perudyptes, Archaeospheniscus, Icadyptes salasi o l'enorme Anthropornis nordenskjoeldi, que feia 170 cm d'alçada i pesava 90 kg. En comparació, el pingüí emperador, el pingüí vivent més gran, només arriba a 122 cm i 37 kg.
Els anseriformes començaren a diversificar-se, amb gèneres com ara Presbyornis, que estava relacionat amb els ànecs i les oques d'avui en dia. L'estudi de la família Presbyornithidae és important per entendre l'evolució de les aus; les espècies que vivien en zones costaneres i que estaven menys especialitzades tendien a extingir-se menys que les espècies que estaven molt adaptades a un ecosistema en concret.
Palaeotis és un estrucioniforme del qual s'han trobat fòssils al jaciment de Messel (Alemanya). És particularment interessant perquè la tesi tradicional sosté que els estrucioniformes tenen els seus orígens a Gondwana, basant-se en la seva difusió actual. El descobriment de Palaeotis a Europa, juntament amb el d'altres estrucioniformes a Nord-amèrica i Mongòlia, posa en dubte aquesta hipòtesi.[19]
A les fosforites del Carcí s'han trobat proves d'una difusió més àmplia dels lloros en temps de l'Eocè. Les restes fòssils del gènere Quercypsitta (fa 37-34 milions d'anys) indiquen que els lloros arribaren en el passat a latituds més septentrionals de les que es troben actualment.[20] Aquests animals, probablement una de les primeres branques de lloros, arribaren a Europa i s'extingiren a tot tardar al Miocè, coincidint amb un refredament del clima global.
Mamífers
[modifica]L'esdeveniment més important en l'evolució dels mamífers durant l'Eocè fou probablement l'evolució dels cetacis. Després que els seus avantpassats abandonessin la vida aquàtica 300 milions d'anys abans, un grup de mamífers relacionat amb els artiodàctils primitius aconseguí efectuar la transició d'un medi terrestre a un medi aquàtic. Mentre els cetacis colonitzaven els medis marins, aparegueren els ratpenats, que es convertiren en els únics mamífers (i un dels pocs grups de vertebrats) capaços de volar.[21] La seva morfologia era extremament similar a l'actual.[22] Es creu que els ratpenats evolucionaren a partir de petits mamífers arborícoles que saltaven d'un arbre a l'altre, desenvolupant en primera instància membranes per planar i finalment ales.[23] Tanmateix, no s'ha descobert cap fòssil que representi un estadi intermedi d'aquesta evolució.[22]
Aquest procés començà amb els pakicètids de l'Eocè inferior i mitjà del Pakistan. Es tractava d'animals carnívors terrestres, però la configuració dels ossos de les orelles i de la seva dentadura demostra que representen el primer pas en l'evolució de les balenes. Uns quants milions d'anys més tard, criatures com Ambulocetus ja tenien un estil de vida amfibi, car les seves potes posteriors estaven més ben adaptades per nedar que per caminar a la terra ferma.[24] Els protocètids són un pas posterior en l'evolució dels cetacis i és possible que disposessin d'una aleta caudal com la dels cetacis actuals.[25]
Els primers cetacis completament marins aparegueren fa uns 45 milions d'anys. Els basilosàurids, que incloïen gèneres com Basilosaurus o Dorudon, tenien una anatomia molt similar a la de les balenes actuals. Tanmateix, els seus cervells estaven menys desenvolupats i no tenien el meló típic dels odontocets actuals. Calgué esperar fins a l'Eocè molt tardà per veure l'aparició de les primeres balenes dentades.

Aquest període també veié l'aparició dels sirenis, un ordre de mamífers marins que inclou els manatís i els dugongs. Els seus representants més primitius eren quadrúpedes amfibis dels gèneres Prorastomus i Pezosiren, dels quals s'han trobat fòssils a Jamaica. Tot i això, es creu que l'origen de l'ordre se situa a Àfrica i que els sirenis estarien emparentats amb els proboscidis, dins el clade dels tetiteris. A la fi de l'Eocè mitjà i durant l'Eocè superior un bon nombre de representants dels sirenis ja eren presents a les aigües de l'oceà de Tetis, a Europa (sobretot Espanya, Itàlia i Hongria) però sobretot a la costa nord-africana (Egipte i Líbia) i Àsia (Índia i Pakistan).
Els ungulats també continuaren evolucionant durant l'Eocè. Els artiodàctils aparegueren a principis d'aquesta època, fa uns cinquanta-quatre milions d'anys, i a la fi de l'Eocè ja s'havien diversificat en els tres subordres actuals: Tylopoda (camells), Suina (porcs) i Ruminantia (ovelles, cabres i vaques). El gran desenvolupament dels perissodàctils, que els desterraren a hàbitats menys pròspers, i l'extensió de l'herba a l'Eocè condicionaren el desenvolupament del particular aparell digestiu que tenen els artiodàctils i que més endavant els ajudaria a desbancar els perissodàctils com a herbívors dominants.[26]
Els carnívors dominants de l'Eocè eren els creodonts,[27] animals semblants als carnívors però que no hi estan directament relacionats. Igual que els carnívors, es caracteritzaven per la presència de dents carnisseres, però en els creodonts, ambdues carnassials eren molars (en els carnívors, són una molar i una premolar).[28] Els creodonts inclouen alguns dels mamífers depredadors terrestres més grans que mai hagin existit, com ara Andrewsarchus, que feia uns tres metres i mig de llargada, gairebé dos metres d'alçada i pesava aproximadament 250 kg.[29] Tanmateix, la seva mida no els serví per imposar-se a llarg termini, foren superats pels carnívors i acabaren extingint-se al Miocè. Això es deu a un nombre de factors. D'una banda, la seva articulació lumbosacra no estava tan adaptada per córrer com la dels carnívors, i la seva condició de plantígrads també els feia menys eficients a l'hora de córrer.[30] D'altra banda, els creodonts tenien una dentadura diferent que els feia carnívors estrictes, mentre que els miàcids i la majoria de carnívors vivents (tret dels felins, que tenen molars vestigials) encara posseïen dents adaptades per menjar altres tipus d'aliment.[28] Dissopsalis, l'últim creodont que apareix en el registre fòssil, s'extingí fa vuit milions d'anys.
Peixos
[modifica]
L'Eocè fou una època en què els taurons lamniformes es diversificaren notablement. Les guilles[31] i els taurons follet[31] foren alguns dels taurons que aparegueren per primer cop. Una de les espècies més destacables fou Otodus obliquus, un tauró que havia aparegut al Paleocè, podia fer fins a nou metres[32] i que s'alimentava de mamífers marins, peixos i altres taurons. Molts paleontòlegs creuen que O. obliquus és l'avantpassat de la família de taurons del gènere Charcharocles i, per tant, que està relacionat amb el tauró depredador més gran que mai hagi existit, C. megalodon.[33] D'altres, en canvi, relacionen Otodus amb el tauró blanc, però cada cop hi ha més evidència que Otodus és molt més proper C. megalodon.[34]
Un altre peix destacable fou Enchodus, un depredador relacionat amb els salmons. Enchodus tenia una mena d'ullals a la part anterior dels maxil·lars superior i inferior i als ossos palatins que li han guanyat el curiós sobrenom d'areng dent de sabre entre els paleontòlegs. Tot i ser un depredador, gran part dels seus fòssils s'han trobat dins els estómacs d'altres depredadors més grans, com els mosasaures, plesiosaures o l'au marina Baptornis.[35]
Rèptils
[modifica]La serp més gran coneguda visqué a la darreria de l'Eocè. Gigantophis garstini podria haver superat els deu metres de longitud, mentre que les serps més grans vivents, les anacondes, mesuren al voltant de set metres de llargària. Aquesta serp, que visqué fa quaranta milions d'anys en el que avui en dia és el Sàhara meridional, podria haver-se alimentat de proboscidis basals.[36] Una altra serp de grans dimensions d'aquesta època fou Palaeophis, una serp marina. Al principi, es calculà que la seva longitud era d'entre trenta i quaranta metres, quelcom d'extraordinari per un ofidi, però estimacions més recents ofereixen una longitud d'uns nou metres.[37]
Artròpodes
[modifica]
En aquesta època, el fet més rellevant en relació amb els artròpodes fou l'expansió de les formigues. Mentre que al Cretaci només n'hi havia unes quantes espècies esteses arreu de Lauràsia i només representaven l'1% dels insectes, al final del Paleocè començaren una radiació adaptativa que es prolongà durant l'Eocè i les situà com a insectes dominants abans de la fi de l'època. El seu èxit fou tal que el 90% de les espècies de formiga vivents a l'Eocè encara existeixen en l'actualitat.[38]
S'han trobat fòssils de Formicium, un gènere de formiga, al jaciment de Messel (Alemanya). És conegut únicament a partir de restes de mascles i de reines, inclou cinc espècies. L'envergadura de les reines era d'entre tretze i quinze centímetres, superior a la de qualsevol altra formiga vivent o extinta. També s'han trobat fòssils d'aquest gènere a Gran Bretanya i als Estats Units, però en aquests casos només es tracta d'ales aïllades.
Flora
[modifica]
A principis de l'Eocè, les altes temperatures i els oceans càlids crearen un ambient humit i calorós, amb boscs que s'estenien de pol a pol. Excepte els deserts més secs, la Terra devia estar completament coberta de boscs.[7]
Els boscs polars eren bastant extensos. S'han trobat fòssils i fins i tot restes preservades d'arbres com ara les cupressàcies o el gènere Metasequoia a Ellesmere Island, a l'àrtic canadenc. Les restes preservades que s'hi han trobat no són fòssils, sinó fragments originals que es conservaren en aigües pobres en oxigen als boscs pantanosos de l'Eocè i després foren enterrades abans que es poguessin descompondre.[39] Fins i tot en aquells temps, Ellesmere Island es trobava només uns quants graus més al sud que en l'actualitat. També s'han trobat fòssils d'arbres subtropicals i fins i tot tropicals de l'Eocè a Groenlàndia i Alaska. Les jungles arribaven fins a latituds tan septentrionals com el nord-oest dels Estats Units i Europa.[7]
A principis de l'època, creixien palmeres a Alaska i al nord d'Europa, tot i que esdevingueren menys abundants a mesura que el clima es refredava. Les Metasequoia també estaven àmpliament esteses.
El refredament començà a mitjans de l'època, i a finals de l'Eocè l'interior dels continents havia començat a assecar-se, i en algunes zones els boscs es reduïren considerablement. L'herba, que havia aparegut feia relativament poc, romania confinada a les ribes dels rius i encara no s'havia estès a les planes i les sabanes.[40]
Amb el refredament vingueren els canvis estacionals. Els arbres caducifolis, més adaptats als grans canvis de temperatura, començaren a imposar-se a les espècies perennes tropicals. A finals de l'època, els boscs caducifolis cobrien grans regions dels continents septentrionals, incloent-hi Nord-amèrica, Euràsia i l'Àrtic, mentre que les jungles només resistien a Sud-amèrica, l'Índia i Austràlia.
L'Antàrtida, que començà l'Eocè amb boscs temperats-subtropicals, es refredà significativament a mesura que passava el temps. La flora tropical de temperatures altes desaparegué, i a principis de l'Oligocè, el continent antàrtic albergava boscs caducifolis i grans regions de tundra.
Grande Coupure
[modifica]
La Grande Coupure, o ‘gran trencament’ en la continuïtat de la població de mamífers d'Europa fa aproximadament 33,5 milions d'anys, marca la fi de l'última fase dels canvis de l'Eocè, del Priabonià i l'arribada a Europa d'immigrants asiàtics. Es caracteritza per grans extincions i especiació al·lopàtrica en petites poblacions aïllades d'espècies primitives. El paleontòleg suís Hans Georg Stehlin encunyà aquest nom el 1910[41] per referir-se al canvi dràstic en els mamífers europeus, que situà al límit Eocè-Oligocè. Un canvi similar en la fauna asiàtica ha rebut el nom de «Remodelat Mongol».
La Grande Coupure marca un trencament entre la fauna endèmica europea anterior al canvi i la fauna mixta, amb un fort component asiàtic, de després del canvi. J. J. Hooker i el seu equip resumiren així el trencament:
| « | A la fauna d'abans de la Grande Coupure són dominants la família de perissodàctils Palaeotheriidae (parents distants dels cavalls); sis famílies d'artiodàctils (Anoplotheriidae, Xiphodontidae, Choeropotamidae, Cebochoeridae, Dichobunidae i Amphimerycidae); la família de rosegadors Pseudosciuridae; les famílies de primats Omomyidae i Adapidae; i la família de soricomorfs Nyctitheriidae. | » |
| « | La fauna de després de la Grande Coupure inclou els autèntics rinoceronts (Rhinocerotidae); tres famílies d'artiodàctils (Entelodontidae, Anthracotheriidae i Gelocidae), relacionades respectivament amb els porcs, hipopòtams i remugants; les famílies de rosegadors Cricetidae (hàmsters) i Castoridae (castors); i la família d'insectívors Erinaceidae (eriçons). Els gèneres Palaeotherium i Anoplotherium, així com les famílies Xiphodontidae i Amphimerycidae, desaparegueren completament. | » |
| « | Només la família de marsupials Herpetotheriidae; la família d'artiodàctils Cainotheriidae; i les famílies de rosegadors Theridomyidae i Gliridae (lirons) sortiren indemnes d'aquest trencament. | » |
| — [42] | ||
Una de les causes principals d'aquest fet sembla haver estat el tancament de l'estret de Turgai, que uní Europa i Àsia i posà fi a l'aïllament geogràfic d'Europa, permetent així migracions massives d'espècies entre ambdós continents. A més, l'obertura del passatge de Drake accentuà el corrent circumpolar antàrtic, iniciant així un refredament progressiu que causà la formació d'un casquet de glaç a l'Antàrtida,[43] així com la formació d'una capa d'aigua freda sobre el fons oceànic. La formació de casquets provocà una important baixada del nivell del mar i accentuà l'albedo de la Terra, reflectint la radiació solar i causant un gran descens de les temperatures. La capa d'aigua freda provocà que moltes de les espècies que vivien en aigües càlides desapareguessin, deixant una fauna molt poc diversa. El canvi climàtic que estava tenint lloc seria la primera fase de les primeres glaciacions polars.
Els mamífers patiren una gran renovació. Els tàxons de mamífers endèmics europeus foren substituïts per immigrants asiàtics, extingint-se així més de la meitat dels mamífers europeus, i afectant d'aquesta manera la resta de la flora i fauna. Els mol·luscs també patiren una gran renovació, especialment a la costa oest dels Estats Units i la plana costanera del nord del Golf de Mèxic.[44]
Una característica de la Grande Coupure era l'extinció aparent de tots els primats europeus; la descoberta recent d'un omòmid de la mida d'un ratolí de principis de l'Oligocè,[45] reflectint les millors possibilitats de supervivència dels mamífers petits, refutà aquesta hipòtesi.
Impactes de meteorit
[modifica]Algunes teories assenyalen l'impacte d'un meteorit sobre Sibèria com a causa principal de la Grande Coupure, a causa d'anomalies trobades en les traces d'iridi, un element associat als impactes de meteorits. L'impacte hauria tingut lloc fa aproximadament trenta-quatre milions d'anys.[46][47] Inicialment es considerà un únic esdeveniment, però més endavant es plantejà un patró d'extinció en massa esglaonada, amb diversos impactes de cometes o meteorits.[48][49][50] Tanmateix, aquest model d'extinció esglaonada fou posteriorment descartat quan es feu evident que els impactes foren anteriors a les extincions,[51] cosa que fou confirmada més endavant gràcies a l'estudi de múltiples talls i sondejos oceànics.[52] Aquests sondejos han estat datats amb gran precisió mitjançant l'anàlisi dels foraminífers plànctonics, arribant a la conclusió que la seva extinció no fou causada pels impactes.[53]
El descobriment posterior de quarsos amb metamorfismes de xoc i espinel·les riques en níquel,[54][55][56] així com el descobriment de diversos cràters a Sibèria i Nord-amèrica, han confirmat que fa aproximadament 35,6 milions d'anys tres grans meteorits impactaren contra la Terra.[57] Alguns autors han suggerit que aquests impactes acceleraren el refredament global,[58][59] tot i que les dades isotòpiques no donen suport a aquesta teoria.[43]
Jaciments
[modifica]Aquests són alguns jaciments destacats de l'Eocè.
Wadi al-Hitan
[modifica]Wadi al-Hitan (وادي الحيتان, 'vall de les balenes' en àrab) és una regió del desert occidental d'Egipte que conté importants restes fòssils de cetacis primitius. Aquestes restes representen un dels principals registres de la història de l'evolució de les espècies: la transformació d'animals terrestres en animals aquàtics soferta per les balenes. Els fòssils trobats a Wadi al-Hitan permeten saber que, en temps de l'Eocè, el que actualment és el desert del Sàhara era un mar som de l'oceà de Tetis.
Els fòssils complets o gairebé complets de Zeuglodon són les troballes més importants que s'han fet a la vall, cosa que li ha guanyat el sobrenom de "Vall dels Zeuglodon". Dorudon és un altre cetaci prehistòric trobat a Wadi al-Hitan. El 2005 la vall fou declarada Patrimoni de la Humanitat per la UNESCO.
Formació del Green River
[modifica]
La Formació del Green River és una formació geològica que s'estén al nord de Utah, l'oest de Colorado i el sud-oest de Wyoming (Estats Units). Conté dues zones distintes de fangs llimosos molt fins, que destaquen per la seva conservació d'una gran diversitat de fòssils complets i detallats. La zona més productiva, anomenada zona de 18 polzades, consisteix en una sèrie de varves i capes laminades de fang que contenen molts peixos i altres fòssils i representa uns quatre mil anys de dipòsits. La segona zona fossilífera és una capa no laminada d'una mica menys de dos metres de gruix que també conté fòssils detallats, però és més difícil extreure'ls perquè no es compon de làmines separables.[60]
Seymour Island
[modifica]Seymour Island és una de les setze grans illes al voltant de la punta de la península antàrtica.
El capità noruec Carl Anton Larsen descobrí fòssils a l'illa el 1882, en el seu primer viatge a Seymour Island a bord del vaixell Jason. Des d'aleshores, l'illa ha estat objecte d'investigacions paleontològiques.
S'hi ha estudiat intensivament el refredament del clima de l'Eocè, un procés que culminà en l'inici de la glaciació de l'Antàrtida. Estudis del carbonat de diversos punts de l'oceà Antàrtic suggereixen que, en lloc de tractar-se d'una baixada monotònica de la temperatura durant aquesta època, a l'Eocè mitjà hi hagué un breu episodi d'escalfament. També s'hi han estudiat molts fòssils, com ara pingüins extints, alguns bivalves, altres animals i algunes plantes.[61]
Argila de Londres
[modifica]L'Argila de Londres és un dipòsit marí conegut pels seus fòssils. És el jaciment més important de l'Ipresià del sud d'Anglaterra, i l'únic lloc d'Europa on es troba una varietat de fòssils vegetals de l'Eocè inferior.
L'argila fou dipositada en un mar que arribava als dos-cents metres de profunditat a la seva regió oriental. S'han trobat fins a cinc cicles de deposició (el que representa una transgressió seguida per una reducció de la profunditat del mar), sobretot a la regió occidental, més soma. Cada cicle comença amb material bast (incloent-hi còdols de sílex arrodonits) i acaba amb una argila cada vegada més sorrenca.[62]

Els fòssils d'animals inclouen bivalves, gastròpodes, nàutils, cucs tubulars, estrelles de potes espinoses, crancs, llagostes, peixos (incloent-hi dents de tauró i de rajada), rèptils (especialment tortugues), i rars exemplars d'ocells. També s'hi han trobat alguns fòssils de mamífers.
Hi són abundants els fòssils de plantes, incloent-hi fruits i llavors. Fa gairebé tres segles que es recullen fòssils vegetals de l'Argila de Londres, i se n'han descrit unes 350 espècies. Això fa que la flora de l'argila de Londres sigui una de les més diverses en fruits i llavors fòssils.[63]
Jaciment de Messel
[modifica]El jaciment de Messel (Hessen, Alemanya) és una antiga pedrera de pissarra bituminosa que comprèn la mostra més ben conservada de la fauna i flora geiseltalianes que es coneix. En molts altres jaciments, trobar esquelets parcials ja és molt, però a Messel hi ha nombrosos casos de conservació completa, que en alguns casos significa que fins i tot han quedat conservats el pelatge, les plomes o les marques de la pell. La diversitat d'espècies també és molt sorprenent (gràcies, en part, a les erupcions de gas). Entre d'altres, al jaciment s'hi han trobat:[64]
- Més de deu mil peixos fossilitzats pertanyents a diverses espècies
- Milers d'insectes aquàtics i terrestres, alguns dels quals encara conserven la coloració
- Una plètora de petits mamífers que inclouen cavalls nans, grans ratolins, primats, opòssums, armadillos, parents del porc formiguer i ratpenats.
- Nombrosos ocells, especialment espècies depredadores
- Cocodrils, granotes, tortugues, salamandres i altres rèptils i amfibis
- Més de trenta restes de plantes, com ara fulles de palmera, fruits, pol·len, fusta, nous i vinyes
Notes
[modifica]- ↑ En els parlars orientals.
- ↑ En els parlars occidentals.
Referències
[modifica]- ↑ Riba i Arderiu i Reguant i Serra, 1986, «Eocè».
- ↑ Rohde, Robert A. «Ypresian ICS Stage». GeoWhen Database. International Commission on Stratigraphy, 2005. Arxivat de l'original el 2008-02-04. [Consulta: 1r març 2008].
- ↑ Rohde, Robert A. «Lutetian ICS Stage». GeoWhen Database. International Commission on Stratigraphy, 2005. [Consulta: 1r març 2008].
- ↑ Rohde, Robert A. «Bartonian ICS Stage». GeoWhen Database. International Commission on Stratigraphy, 2005. Arxivat de l'original el 2008-02-04. [Consulta: 1r març 2008].
- ↑ Rohde, Robert A. «Priabonian ICS Stage». GeoWhen Database. International Commission on Stratigraphy, 2005. [Consulta: 1r març 2008].
- ↑ Stanley, Steven M. Earth System History. Nova York: W.H. Freeman and Company, 1999. ISBN 0716728826 [Consulta: 5 abril 2008].
- ↑ 7,0 7,1 7,2 Agaric. «Earth's Eocene Epoch». Associated Content, 2007. Arxivat de l'original el 2013-01-02. [Consulta: 1r març 2008].
- ↑ http://pubs.giss.nasa.gov/docs/2003/2003_Schmidt_Shindell.pdf Arxivat 2011-10-20 a Wayback Machine.
- ↑ White, Kasey. «Clues to global warming». UC Santa Cruz, 2003. Arxivat de l'original el 2008-07-06. [Consulta: 7 març 2008].
- ↑ 10,0 10,1 Pagani, M.; Pedentchouk, N., Huber, M., Sluijs, A., Schouten, S., Brinkhuis, H., Sinninghe Damsté, J.S., Dickens, G.R., Others, «Arctic hydrology during global warming at the Palaeocene/Eocene thermal maximum». Nature, 442, 7103, 2006, pàg. 671-675.
- ↑ Tim Appenzeller «Great green north». National Geographic, 5-2005. Arxivat de l'original el 2007-06-01 [Consulta: 1r març 2008]. Arxivat 2007-06-01 a Wayback Machine.
- ↑ Doug MacDougall. Frozen Earth: The Once and Future Story of Ice Ages. University of California Press, 2004. ISBN 0-520-24824-4.
- ↑ Kieran Mulvaney. At the Ends of the Earth: A History of the Polar Regions. Washington, D.C.: Island Press, 2001. ISBN 1-55963-908-3.
- ↑ «Tectonics of the Eocene». UC Museum of Paleontology, 1999. [Consulta: 7 març 2008].
- ↑ «Ring of Fire». United States Geological Survey, 2008. Arxivat de l'original el 2006-08-28. [Consulta: 7 març 2008].
- ↑ Zhu, Bin; William S. F. Kidd, David B. Rowley, Brian S. Currie, i Naseer Shafique «Age of Initiation of the India-Asia collision in the East-Central Himalaya». Journal of Geology. Arxivat de l'original el 2005-12-21 [Consulta: 2 març 2008].
- ↑ Husson, Laurent; Jean-Louis Mugnier, Pascal Leturmy i Gérard Vidal «Kinetics and Sedimentary Balance of the Subhimalayan Zone, West Nepal». Thrust Tectonics and Hydrocarbon Systems, 82, 2004, pàg. 115-130. Arxivat de l'original el 2007-04-18 [Consulta: 4 març 2008]. Arxivat 2007-04-18 a Wayback Machine.
- ↑ Haines, Tim. «New Dawn». A: Walking with Beasts. Londres: BBC Books, 2001.
- ↑ Chatterjee, Sankar «The Rise of Birds». Johns Hopkins University Press, 1997.
- ↑ Mourier-Chauviré, Cécile «Une nouvelle famille de Perroquets (Aves, Psittaciformes) dans l'Eocène supérieur des Phosphorites du Quercy». Geobios, Mémoire Spécial, 14, 1992, pàg. 169-177.
- ↑ Bob Holmes: How bats made the leap from gliding to flying. New Scientist: volum 199, número 2664 (12 de juliol del 2008). Pàgina 16.
- ↑ 22,0 22,1 Laurent, Arthur & Lemaire, Michel (1999). Les chauves-souris, maîtresses de la nuit. París, Editorial Delachaux-Niestlé. ISBN 2-603-01147-2
- ↑ David Attenborough, BBC, 2002. The Life of Mammals. Episodi 2, Insect Hunters.
- ↑ Sutera, Raymond «The Origin of Whales and the Power of Independent Evidence». , 2000.
- ↑ Clementz, Marc T.; Goswami, A., Gingerich, P., Koch, P. «Isotopic records from early whales and sea cows: contrasting patterns of ecological transition». Journal of Vertebrate Paleontology, 26:2, 2006, pàg. 355–370.
- ↑ Janis, Christine & Jarman, Peter. Macdonald, D.. The Encyclopedia of Mammals. Nova York: Facts on File, 1984, p. 498-499. ISBN 0-87196-871-1.
- ↑ David Lambert and the Diagram Group. The Field Guide to Prehistoric Life. Nova York: Facts on File Publications, 1985. ISBN 0-8160-1125-7
- ↑ 28,0 28,1 David McDonald. «The Carnassial Connection». A: The Velvet Claw: A Natural History of the Carnivores. BBC Books, 1993. ISBN 0563208449.
- ↑ Haines, Tim. «Land of Giants». A: Walking with Beasts. Londres: BBC Books, 2001 [Consulta: 30 desembre 2007].
- ↑ «The Elements of Geology». Globusz. Arxivat de l'original el 2008-03-09. [Consulta: 11 març 2008].
- ↑ 31,0 31,1 Sepkoski, Jack «A compendium of fossil marine animal genera (Chondrichthyes entry)». Bulletins of American Paleontology, 364, 2002, pàg. 560. Arxivat de l'original el 2012-05-10 [Consulta: 1r setembre 2008].
- ↑ Renz, Mark. Megalodon: Hunting the Hunter. PaleoPress, 2002, p. 26-30.
- ↑ Andres, Les. Evolutionary lineage of Sharks.
- ↑ Carcharocles. elasmo, 2005.
- ↑ Everhart, Mike. «Enchodus sp. - The Sabre-Toothed Fish of the Cretaceous». Oceans of Kansas, 2007. [Consulta: 23 agost 2007].
- ↑ New Scientist, nº 2473, pàg. 17. 13 novembre 2004.
- ↑ Parmley, Dennis; Melanie DeVore «Palaeopheid Snakes from the Late Eocene Hardie Mine Local Fauna of Central Georgia». Southeastern Naturalist, 4:4, 2005, pàg. 703-722.
- ↑ D. Grimaldi & D. Agosti «A formicine in New Jersey Cretaceous amber (Hymenoptera: Formicidae) and early evolution of the ants». Proceedings of the National Academy of Sciences, 97, 2001, pàg. 13678–13683.
- ↑ http://www.fossilmuseum.net/plantfossils/plfossil8/Metasequoia.htm
- ↑ «Desert Grasses». Arizona-Sonora Desert Museum, 2007. [Consulta: 11 març 2008].
- ↑ H.G. Stehlen, 1910. "Remarques sur les faunules de Mammifères des couches eocenes et oligocenes du Bassin de Paris," in Bulletin de la Société Géologique de France, 4'.9, pp 488-520.
- ↑ Hooker et al. 2004
- ↑ 43,0 43,1 Livermore, Roy; Nankivell, Adrian; Eagles, Graeme; Morris, Peter «Paleogene opening of Drake Passage». Earth and Planetary Science Letters, 236, 1-2, 2005. pàg. 459-470.
- ↑ Ivany L.C., Patterson W.P., Lohmann K.C. «Cooler winters as a possible cause of mass extinctions at the Eocene/Oligocene boundary». Nature, 407, 2000. pàg. 887-890.
- ↑ Meike Kohler i Salvador Moya-Sola, "A finding of Oligocene primates on the European continent," a Proceedings of the National Academy of Sciences of the United States 96.25 (7 desembre 1999), pp 14664-14667
- ↑ name="Ganapathy1982">Ganapathy, R. «A Major Meteorite Impact on the Earth 65 Million Years Ago: Evidence from the Cretaceous-Tertiary Boundary Clay». Science, 4459, 216, 1982, pàg. 885-886.
- ↑ Álvarez, W., Asaro, F., Michel, H.V., Álvarez, L.W. «Iridium anomaly approximately synchronous with terminal Eocene extinctions». Science, 216, 4548, 1982, pàg. 886-888.
- ↑ Hut, P., Álvarez, W., Elder, W.P., Hansen, T., Kauffman, E.G., Keller, G., Shoemaker, E.M., Weissman, P.R. «Comet showers as a cause of mass extinctions». Nature, 329, 1987, pàg. 118-126.
- ↑ Keller, G. «Stepwise mass extinctions and impact events: Late Eocene and early Oligocene». Marine Micropaleontology, 13, 1986, pàg. 267-293.
- ↑ Kauffman, E.G. «The dynamics of marine stepwise mass extinction». Revista Española de Paleontología, Extraordinario, 1988, pàg. 57-71.
- ↑ Keller, G., D'Hondt, S. L., Orth, C. J., Gilmore, J. S., Oliver, P. Q., Shoemaker, E. M., Molina, E. «Late Eocene impact microspherules - Stratigraphy, age and geochemistry». Meteoritics, 22, Març, 1987, pàg. 25-60.
- ↑ Molina, E.; Gonzalvo, C., Keller, G «The Eocene-Oligocene planktic foraminiferal transition: extinctions, impacts and hiatuses». Geological Magazine, vol. 130, núm. 4, 1993, p. 483-499.
- ↑ Gonzalvo, C. & Molina, E. «Bioestratigrafía y cronoestratigrafía del tránsito Eoceno-Oligoceno en Torre Cardela y Massignano (Italia)». Revista Española de Paleontología, 7, 2, 1992, pàg. 109-126.
- ↑ Clymer, Aron K., Bice, David M., Montanari, Alessandro «Shocked quartz from the late Eocene: Impact evidence from Massignano, Italy». Geology, 24, 6, 1996, pàg. 483-486.
- ↑ Pierrard, O., Robin, E., Rocchia, R., Montanari, A. «Extraterrestrial Ni-rich spinel in upper Eocene sediments from Massignano, Italy». Geology, 26, 4, 1998, pàg. 307-310.
- ↑ Molina, E., Gonzalvo, C., Ortiz, S., Cruz, L.E. «Foraminiferal turnover across the Eocene-Oligocene transition at Fuente Caldera, southern Spain: no cause-effect relationship between meteorite impacts and extinctions». Marine Micropaleontology, 58, 2006, pàg. 270-286.
- ↑ Poag, C.W., Mankinen, E., Norris, R.D.. «Late Eocene Impacts: Geologic Record, Correlation and Paleoenvironmental Consequences». A: From Greenhouse to Icehouse. Nova York: Columbia University Press, 2003, p. 495-510. ISBN 0-231-12716-2.
- ↑ Wonhof, H.B., Smit, J., Brinkhuis, H., Montanari, A., Nederbragt, A.J. «Global cooling accelerated by early late Eocene impacts». Geology, 28, 8, 2000, pàg. 687-690.
- ↑ Bodiselitsch, B; Montanari, A., Koeberl, C., Coccioni, R. «Delayed climate cooling in the Late Eocene caused by multiple impacts: high-resolution geochemical studies at Massignano». Earth and Planetary Science Letters, 223, 3-4, 2004, pàg. 283-302.
- ↑ http://www.ucmp.berkeley.edu/tertiary/eoc/greenriver.html
- ↑ «Middle Eocene Warming On Seymour Island, Antarctica: Continental Shelf Paleotemperatures Recorded In Molluscan Carbonates». Arxivat de l'original el 2008-08-21. [Consulta: 11 març 2008].
- ↑ Sumbler, M.G.. London and the Thames Valley (en anglès). 4a edició. British Geological Survey, 1996 (British Regional Geology series). ISBN 0-11-884522-5.
- ↑ Collinson, Margaret E. The Palaeontological Association. Fossil plants of the London Clay (en anglès). Wiley-Blackwell, 1983. ISBN 978-0901702265.
- ↑ Grube Messel
Bibliografia
[modifica]- Riba i Arderiu, O.; Reguant i Serra, S. Una taula dels temps geològics. Institut d'Estudis Catalans, 1986. ISBN 9788472830776.
Enllaços externs
[modifica]- Eocè Arxivat 2009-06-11 a Wayback Machine.
- Paleomapa de l'Eocè
- Eocene life Arxivat 2004-10-10 a Wayback Machine. (anglès)
{kind=link}
|
|
Aquest article conté una o més imatges SVG amb text que pot ser traduït al català. (ajuda) |