
Thalassodromeus
 Reconstrucció de T. sethi (amb un postcrani hipotètic) al Museu Nacional de la Natura i la Ciència de Tòquio | |
| Període | |
|---|---|
| Estat de conservació | |
| Taxonomia | |
| Superregne | Holozoa |
| Regne | Animalia |
| Fílum | Chordata |
| Classe | Reptilia |
| Ordre | Pterosauria |
| Família | Thalassodromidae |
| Gènere | Thalassodromeus Kellner i Campos, 2002 |
| Nomenclatura | |
| Sinònims | |
| Significat | Corredor del mar |
| Espècies | |
| |
| Mida | |
 | |
Thalassodromeus és un gènere de pterosaures de la família dels talassodròmids que visqueren en allò que avui en dia és el Brasil durant el Cretaci inferior, fa aproximadament 100 milions d'anys.
El crani original, descobert el 1983 a la conca de l'Araripe, al nord-est del Brasil, fou excavat peça a peça en moments diferents. El 2002, els paleontòlegs Alexander Kellner i Diógenes de Almeide Campos el designaren com a holotip de l'espècie T. sethi. El nom genèric Thalassodromeus significa 'corredor del mar', en referència al mode d'alimentació que se suposa que tenia, i el nom específic sethi es refereix a Seth, un déu egipci que portava una corona que, segons els científics, recorda la cresta d'aquest pterosaure. Altres investigadors han suggerit que la cresta és més aviat similar a la corona d'Amon. El 2005 es referí l'extrem d'una mandíbula a T. sethi, però el mateix espècimen es convertí en l'holotip del nou gènere Banguela el 2014, abans de tornar a ser classificat en el si de Thalassodromeus com una nova espècie, T. oberlii. Una tercera espècie, T. sebesensis, fou descrita el 2015 a partir d'un presumpte fragment de la cresta que al final resultà ser un tros de closca de tortuga.
Thalassodromeus tenia un dels cranis més grans coneguts entre els pterosaures, d'uns 1,42 m de llarg, amb una de les crestes cranials proporcionalment més grans de qualsevol vertebrat. Tot i que només es coneix el crani, es calcula que l'animal tenia una envergadura de 4,2 a 4,5 m. La cresta era lleugera i anava des de la punta de la mandíbula superior fins a més enllà de la part posterior del crani, acabant en una osca única en forma de V. Les mandíbules eren sense dents i tenien les vores superiors i inferiors esmolades. El seu crani tenia grans finestres nasoantorbitàries (obertura que combinava la finestra antorbitària davant de l'ull amb la fossa nasal òssia), i part del seu paladar era còncau. La mandíbula inferior era semblant a una fulla, i podria haver girat lleugerament cap amunt.
El parent més proper de Thalassodromeus era Tupuxuara; tots dos s'agrupen en un clade que s'ha situat dins dels Tapejaridae (com la subfamília Thalassodrominae) o dins de Neoazhdarchia (com la família Thalassodromidae).
S'han suggerit diverses teories per explicar la funció de la cresta de Thalassodromeus, incloent la termoregulació i la visualització, però probablement tenia més d'una funció. Les crestes dels talassodromins sembla que s'han desenvolupat tard en el creixement (probablement correlacionades amb la maduresa sexual) i poden haver estat sexualment dimòrfiques (difereixen segons el sexe).
Tal com indica el nom del gènere, Thalassodromeus es va proposar originalment per haver-se alimentat com un ocell bec de tisora modern, passant per sobre de la superfície de l'aigua i submergir les seves mandíbules inferiors per atrapar preses. Aquesta idea va ser criticada més tard per manca de proves. Des d'aleshores s'ha trobat que Thalassodromeus tenia una forta musculatura de la mandíbula i podria haver estat capaç de matar i menjar preses relativament grans a terra. Les proporcions de les extremitats de les espècies relacionades indiquen que podria haver-se adaptat per volar a l'interior i hauria estat eficient per moure's a terra. Thalassodromeus és conegut de la Formació Santana, on va conviure amb molts altres tipus de pterosaures, dinosaures i altres animals.
Història i descobriment
[modifica]El primer exemplar conegut d'aquest pterosaure (un ordre extingit de rèptils voladors) es va recollir l'any 1983 prop de la localitat de Santana do Cariri, a la conca d'Araripe, al nord-est del Brasil. Trobat en afloraments de la Formació Santana, va ser recollit durant un llarg període en diverses peces. L'exemplar (catalogat com a DGM 1476-R al Museu de Ciências da Terra) es va conservar en un nòdul calcari, i consta d'un crani tridimensional gairebé complet (els ossos de pterosaure solen ser fòssils de compressió aplanats), mancant dos segments de la part inferior del crani i la mandíbula i la part davantera de la mandíbula inferior. La regió jugal esquerra i la rama mandibular dreta (la meitat de la mandíbula) s'empenyen lleugerament cap a dins. El crani es va informar per primera vegada en un llibre italià de 1984, i va ser descrit i imaginat preliminarment el 1990 pels paleontòlegs Alexander Kellner i Diògenes de Almeida Campos. Tot i que les peces de crani s'havien repartit entre museus d'Amèrica del Sud i del Nord, es van reunir abans del 2002.[2][3][4][5][6]
L'any 2002, Kellner i Campos van descriure i anomenar el nou gènere i espècie Thalassodromeus sethi, essent el crani DGM 1476-R l'exemplar holotip. El nom genèric deriva de les paraules del grec antic θάλασσα (thálassa, «mar») i δρομεύς (dromeús, «corredor»), que significa «corredor del mar» en referència al suposat comportament d'alimentació piscívora de l'animal. El nom específic fa referència al déu egipci Seth.[2][4] L'exemplar no estava completament preparat en el moment d'aquesta descripció preliminar.[7] Els descriptors originals van triar el nom «sethi» perquè la cresta d'aquest pterosaure suposadament recordava la corona que portava Seth, però els paleontòlegs André Jacques Veldmeijer, Marco Signore i Hanneke J. M. Meijer van assenyalar el 2005 que la corona (amb les seves dues plomes altes) normalment el portava el déu Amon (o Amon-Ra) i les seves manifestacions, no Seth.[7][8]
-
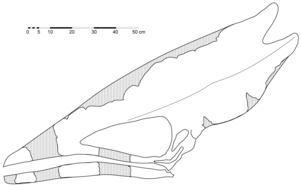 Diagrama del crani holotip de T. sethi (les línies ombrejades representen les parts que falten), basat en la seva interpretació inicial
Diagrama del crani holotip de T. sethi (les línies ombrejades representen les parts que falten), basat en la seva interpretació inicial -
 Reconstrucció del crani de T. sethi, que mostra la possible extensió de la cresta no danyada i les mandíbules cap avall proposada per Headden, però no recolzada per Pêgas i els seus col·legues
Reconstrucció del crani de T. sethi, que mostra la possible extensió de la cresta no danyada i les mandíbules cap avall proposada per Headden, però no recolzada per Pêgas i els seus col·legues -
 Encara que T. sethi va rebre el nom del déu Seth per tenir una cresta semblant a la seva corona, Seth no portava aquesta corona; la cresta era semblant a la corona d'Amon (esquerra)
Encara que T. sethi va rebre el nom del déu Seth per tenir una cresta semblant a la seva corona, Seth no portava aquesta corona; la cresta era semblant a la corona d'Amon (esquerra)



L'any 2006, els paleontòlegs David M. Martill i Darren Naish van suggerir que Thalassodromeus era un sinònim menor del gènere relacionat Tupuxuara, que va ser nomenat per Kellner i Campos l'any 1988 a partir de fòssils de la mateixa formació. Segons Martill i Naish, les diferències entre aquests gèneres (incloent dues espècies de Tupuxuara, T. longicristatus i T. leonardii) es deuen a l'ontogènia (canvis durant el creixement) i a la compressió dels fòssils; Thalassodromeus era simplement un individu més gran, més gran i millor conservat.[9] Aquesta idea va ser rebutjada per Kellner i Campos el 2007, que van assenyalar que aquestes espècies tenien diferències en característiques diferents de les seves crestes. També van assenyalar que un exemplar de Tupuxuara tenia un crani més gran que Thalassodromeus (mesurat des de la punta de la premaxil·la fins a la part posterior de l'os escatós), malgrat l'afirmació de Martill i Naish que aquest últim era un individu més gran.[10] L'opinió de Kellner i Campos ha estat acceptada des d'aleshores per altres investigadors, inclosos Martill i Naish.[7][11]
Veldmeijer i els seus col·legues van assignar la part frontal d'una mandíbula recollida de la mateixa formació a T. sethi l'any 2005. Van concloure que, tot i que els dos exemplars diferien en diversos detalls, les diferències no eren prou significatives com per basar una nova espècie en la mandíbula, i que el nou exemplar va omplir el buit de la reconstrucció del crani T. sethi de Kellner i Campos.[8] Els paleontòlegs Jaime A. Headden i Herbert B. N. Campos van encunyar el nou binomi Banguela oberlii, basant-se en la seva reinterpretació de la punta de la mandíbula com a pertanyent a un membre sense dents de la família Dsungaripteridae, l'any 2014. El nom genèric és portuguès per «desdentat» i el nom específic fa honor al col·leccionista privat Urs Oberli, que havia donat l'exemplar al Naturmuseum St. Gallen (on està catalogat com a NMSG SAO 25109). Headden i Campos van interpretar la punta de la mandíbula inferior de T. sethi cap avall; aquest i altres trets el van distingir de Banguela.[12] En la seva re-descripció del 2018 del crani holotip de T. sethi, més preparat, els paleontòlegs Rodrigo V. Pêgas, Fabiana R. Costa i Kellner van tornar a assignar B. oberlii a Thalassodromeus alhora que el van reconèixer com una espècie diferent i, per tant, van crear la combinació nova T. oberlii. Pêgas i els seus col·legues també van rebutjar la teoria que la mandíbula inferior de T. sethi estava cap avall, i van reinterpretar la peça més davantera de la mandíbula inferior perquè s'havia connectat directament amb la peça posterior (sense escletxa).[7]
L'any 2015 els paleontòlegs Gerald Grellet Tinner i Vlad A. Codrea van anomenar una nova espècie, T. sebesensis, a partir del que van interpretar com a part d'una cresta cranial en una concreció trobada prop del riu Sebeș, a Romania. Els autors van dir que això ampliaria considerablement l'abast en temps i espai del gènere Thalassodromeus, creant una bretxa de 42 milions d'anys entre les espècies sud-americanes més antigues i les espècies europees més joves.[13] El paleontòleg Gareth J. Dyke i un gran equip de col·legues van rebutjar immediatament la identificació de pterosauris del fòssil de T. sebesensis, en lloc d'argumentar que era una part mal identificada d'un plastró (closca inferior) de la tortuga prehistòrica Kallokibotion bajazidi (anomenada el 1923). La idea que el fragment pertanyia a una tortuga havia estat considerada i rebutjada per Grellet-Tinnera i Codrea en la seva descripció original.[14] Grellet-Tinnera i Codrea van negar la identitat de la tortuga suggerida per Dyke i els seus col·legues, assenyalant que aquests investigadors no havien examinat directament el fòssil.[15]
Descripció
[modifica]L'holotip (i únic crani conegut) de Thalassodromeus sethi és un dels cranis de pterosaure més grans que s'han descobert mai. S'estima que tot el crani feia 1,42 m de llarg; els ossos estaven fusionats, indicant l'edat adulta. Basat en pterosaures relacionats, la seva envergadura era de 4,2 a 4,5 m, fent de Thalassodromeus el membre més gran conegut del seu clade, Thalassodromidae. De proporcions similars, el seu crani estava més construït que el del seu parent Tupuxuara.[2][4][7] Tot i que es desconeix l'esquelet postcranial de Thalassodromeus, els seus parents tenien vèrtebres de coll inusualment curtes i en blocs, amb les extremitats anteriors i posteriors ben desenvolupades que eren gairebé iguals en longitud (excepte el dit llarg de l'ala). Les extremitats posteriors eren el 80% de la longitud de les extremitats anteriors, una proporció única entre els pterodactiloïdeus (pterosaures de cua curta).[4] Com a pterosaure, Thalassodromeus estava cobert de picnofibres semblants a pèls i tenia unes membranes alars extenses (que s'estenia amb el dit de l'ala).[16]
El crani de T. sethi tenia un perfil aerodinàmic, sobretot des de la punta del musell fins a la vora frontal de la fenestra nasoantorbitària (obertura que combinava la fenestra antorbitària davant de l'ull amb la fossa nasal òssia). La característica més conspicua del crani era la gran cresta, que anava al llarg de la vora superior des de la punta del musell i més enllà de l'occípit a la part posterior del crani, gairebé duplicant la longitud i l'alçada del crani. Amb l'excepció del pterosaure Tupandactylus imperator (la cresta del qual consistia principalment en teixit tou), T. sethi tenia la cresta cranial proporcionalment més gran de qualsevol vertebrat conegut (75% de la superfície lateral del crani). La cresta estava formada principalment per les premaxil·les (els ossos del musell més davanters), ossos frontals, ossos parietals i part de l'os supraoccipital. Les premaxil·les formaven la major part de la cresta, estenent-se fins a la seva part posterior, i van contactar amb la part frontoparietal de la cresta mitjançant una sutura recta (una característica diferent d'aquesta espècie). La cresta variava d'1 a 10,5 mm de gruix; es va espessir al contacte entre les premaxil·les i la part frontoparietal, i es va anar aprimant cap a la part superior i posterior (excepte la part inferior darrere de l'occípit, on tenia una base gruixuda).[2][4][7]
-
 Còpia del crani holotip de T. sethi, Museu d'Història Natural de Cleveland
Còpia del crani holotip de T. sethi, Museu d'Història Natural de Cleveland -
 Il·lustració d'una parella de T. sethi volant
Il·lustració d'una parella de T. sethi volant


Malgrat la seva mida, la cresta era lleugerament construïda i essencialment buida; algunes zones indiquen signes de pneumatització esquelètica i un sistema trabecular ben desenvolupat que uneix els ossos. La superfície de la cresta tenia un sistema de canals de diferents mides i gruixos, probablement les impressions d'extensos vasos sanguinis.[2][4] Hi havia una petita obertura de 46 mm per sobre de l'òrbita de l'ull, perforant la part basal de la cresta; aquesta característica és desconeguda en altres pterosaures i no sembla que sigui deguda a danys. Els marges de l'obertura són llisos i la vora interior té una fenestració que la connecta amb l'estructura interna de la cresta.[7][10] La part posterior de la cresta acabava en una destacada osca en forma de V, característica única d'aquesta espècie.[2][4] Encara que altres parts de la cresta presenten trencaments en forma de V, la forma de V de l'extrem no sembla haver estat deguda a trencaments; s'hi veuen els marges de l'os, encara tancats per la matriu. La cresta probablement tenia una coberta queratinosa (córnea) i podria haver estat estesa per teixits tous en algunes zones, però es desconeix l'extensió d'aquesta.[2][17]
La mandíbula superior de T. sethi estava composta principalment per premaxil·les i maxil·lars superiors; la sutura que formava la frontera entre aquests ossos no és visible. Com en tots els membres del seu clade, les mandíbules eren edèntules (desdentades). El rostre (musell) feia 650 mm de llarg des de la punta de la premaxil·la fins a l'articulació on l'os quadrat del crani es connectava amb l'os articular de la mandíbula inferior. La part davantera de les premaxil·les tenia vores superiors i inferiors esmolades, exclusives d'aquesta espècie. Com en els gèneres relacionats, la fenestra nasoantorbitària era relativament gran; feia 650 mm de llarg i 200 mm d'alçada, que era el 71% de la longitud del crani (excloent la cresta). L'os lacrimal, que separava l'òrbita de la fenestra nasoantorbitària, era allargat verticalment i més alt que la superfície superior de l'òrbita (en contrast amb la condició observada en pterodactiloïdeus amb fenestra nasoantorbitària més petites). L'òrbita era esvelta i comprimida de davant a darrere en comparació amb els Tupuxuara i els tapejàrids, però semblant a alguns d'ells en ser més de la meitat de l'alçada de la fenestra nasoantorbitària. L'òrbita es va situar més avall que el marge superior de la fenestra nasoantorbitària i, per tant, molt baix del crani. Tot i que els ossos que vorejaven la fenestra temporal inferior (una obertura darrere de l'òrbita) estaven incomplets, sembla que eren allargats i semblants a una escletxa (com a Tupuxuara i Tapejara).[4][7]
La zona del palatal a l'extrem del musell de T. sethi era una cresta esmolada, semblant a la quilla que es veu a la superfície superior de la símfisi mandibular on es connectaven les dues meitats de la mandíbula inferior. Petits foramina (obertures) en forma d'escletxa a les vores laterals inferiors de la carena indiquen que tenia una coberta còrnia en vida, semblant a Tupandactylus. La vora inferior de la zona era una mica corba, cosa que probablement creava un petit buit quan es tancaven les mandíbules. Més enrere, immediatament davant de la fenestra nasoantorbitària, la cresta palatina es va convertir en una quilla forta, roma i convexa. Aquesta convexitat encaixava a la prestatgeria simfisaria a l'extrem frontal de la mandíbula inferior, i s'haurien enganxat fortament quan les mandíbules estaven tancades. La carena palatina acabava en una zona fortament còncava exclusiva d'aquesta espècie. Les fenestrae postpalatinas (obertures darrere de l'os palatí) eren ovalades i molt petites, diferents de les de les espècies relacionades. L'ectopterigoide (os al costat del paladar) tenia costats grans, semblants a plaques, i estava ben desenvolupat en comparació amb les espècies relacionades. L'os supraoccipital, que formava la base més posterior de la cresta cranial, tenia cicatrius musculars a l'extrem superior (probablement corresponen a la fixació dels músculs del coll).[7]
Tot i que la mandíbula inferior de T. sethi està incompleta, la seva longitud total s'estima en 670 mm, el 47% dels quals estava ocupat per la símfisi mandibular. Falta la punta de la mandíbula, però la seva superfície frontal indica que podria haver estat girada lleugerament cap amunt com en T. oberlii (la possible segona espècie de Thalassodromeus, o possiblement un gènere diferent (Banguela) que només es coneix per la punta de la mandíbula). El prestatge de la símfisi (la superfície superior de la símfisi), s'estenia 170 mm i tenia una superfície plana. Vist des de dalt, les vores laterals d'aquesta zona eren altes i formaven un marge nítid. Prop de l'extrem frontal de la símfisi, les vores que formaven els marges es van fer més amples cap a la part davantera del prestatge fins que es van trobar i es van fusionar. Les superfícies superior i inferior de la mandíbula a la part davantera del prestatge eren quillades (la quilla superior més robusta i començant abans de la inferior), cosa que donava a la símfisi una forma de fulla. La quilla inferior es va fer més profunda cap a la part davantera de la mandíbula, donant la impressió que la mandíbula es desviava cap avall; en realitat era recta, excepte per la punta (potser) cap amunt. Les fosses mandibulars (depressions) a la part posterior de la mandíbula superior eren més profundes i amples del que és habitual en els pterodactiloïdeus, creant grans superfícies amb les quals s'articulava la mandíbula inferior. La possible espècie T. oberlii es diferenciava de T. sethi i altres parents perquè la superfície superior de la seva símfisi mandibular era lleugerament més curta que la superfície inferior, i es va distingir encara més de T. sethi perquè la vora superior de la símfisi era molt més esmolada que la més baix. Les dues espècies compartien característiques com la compressió de la símfisi de costat i de dalt a baix, la quilla esmolada a la part davantera superior de la símfisi i el petit solc que recorre la superfície superior del prestatge.[7]
Classificació
[modifica]
La classificació de Thalassodromeus i els seus parents més propers és un dels temes més polèmics pel que fa al seu grup.[4] Kellner i Campos van assignar originalment Thalassodromeus a la família Tapejaridae, basant-se en la seva gran cresta i la seva gran fenestra nasoantorbitària. Dins d'aquest clade, van trobar que era diferent del gènere de cara curta Tapejara, però compartia una quilla al paladar amb Tupuxuara.[2] Kellner va elaborar les relacions entre Tapejaridae el 2004 i va assenyalar que Thalassodromeus i Tupuxuara també compartien una cresta formada principalment per os; la cresta tenia un gran component de teixit tou en altres membres del grup.[17]
Martill i Naish van considerar Tapejaridae un grup parafilètic (antinatural) l'any 2006, i van trobar que Tupuxuara (que va incloure Thalassodromeus en la seva anàlisi) era el tàxon germà de la família Azhdarchidae. Aquest clade (Tupuxuara i Azhdarchidae) havia estat nomenat Neoazhdarchia pel paleontòleg David Unwin l'any 2003, un acord amb el qual van coincidir Martill i Naish. Segons Martill, les característiques que uneixen els membres de Neoazhdarchia inclouen la presència d'un notarium (vèrtebres fusionades a la regió de l'espatlla), la pèrdua de contacte entre el primer i el tercer metacarpià (ossos a la mà) i els musells molt llargs (més del 88%) de la longitud del crani).[9][18] Kellner i Campos van defensar la validesa de Tapejaridae el 2007, dividint-lo en dos clades: Tapejarinae i Thalassodrominae, aquest últim contenint Thalassodromeus (el gènere tipus) i Tupuxuara. Van distingir els talassodromins per les seves elevades fenestrae nasoantorbitàries i la part òssia de les seves crestes començant a la part davantera del crani i continuant més enrere que en altres pterosaures.[10]
La interrelació d'aquests clades dins del clade més gran Azhdarchoidea va romandre en disputa, i el clade que conté Thalassodromeus i Tupuxuara havia rebut diferents noms de diferents investigadors (Thalassodrominae i Tupuxuaridae). El paleontòleg Mark Witton va intentar resoldre el problema de la denominació el 2009, assenyalant que el nom «Tupuxuaridae» (utilitzat per primera vegada en la forma vernacular «tupuxuàrids» pel paleontòleg Lü Junchang i els seus col·legues el 2006) mai s'havia establert vàlidament i Thalassodrominae hauria de ser el nom propi (tot i que es va atorgar un any després). Witton va convertir encara més el nom de subfamília Thalassodrominae en el nom de família Thalassodromidae, i va considerar que el clade formava part de Neoazhdarchia.[11]
Una anàlisi del 2011 del paleontòleg Felipe Pinheiro i els seus col·legues va confirmar l'agrupació dels clades Tapejarinae i Thalassodrominae a la família Tapejaridae, a la qual s'uneixen els Chaoyangopterinae.[19] Un estudi del 2014 del paleontòleg Brian Andres i els seus col·legues va trobar que els talassodromins s'agrupaven amb els Dsungariptèrids, formant el clade Dsungaripteromorpha dins de Neoazhdarchia (definit com el clade més inclusiu que conté Dsungaripterus weii però no Quetzalcoatlus northropi).[20]
Cladograma basat en Pinheiro i els seus col·legues, 2011:[19]
|
Cladograma basat en Andres i els seus col·legues, 2014:[20]
|
Pêgas i els seus col·legues van mantenir Tapejarinae i Thalassodrominae com a part de Tapejaridae el 2018, però van reconèixer que el tema encara era controvertit.[7]
Paleobiologia
[modifica]Funció de la cresta
[modifica]Kellner i Campos van proposar possibles funcions per a la cresta cranial de Thalassodromeus l'any 2002. Van suggerir que la xarxa de vasos sanguinis de la seva gran superfície era coherent amb l'ús per a la termoregulació, que també s'havia suggerit per a les crestes d'alguns dinosaures. Kellner i Campos van pensar que la cresta s'utilitzava per a la refrigeració (permetent a l'animal dissipar l'excés de calor metabòlica per convecció), mentre que la transferència de calor estava controlada per (i depenia) de la xarxa de vasos sanguinis. La capacitat de controlar la seva temperatura corporal hauria ajudat a Thalassodromeus durant una activitat intensa (com la caça), i van suggerir que, en vol, la calor s'hauria dissipat amb més eficàcia si la cresta estigués alineada amb el vent, mentre que el cap estava intencionadament traslladat als costats. Kellner i Campos van plantejar que l'escut podria haver tingut funcions addicionals, com ara l'exhibició; ajudat pel color, podria haver estat utilitzat en el reconeixement d'espècies, i també podria haver estat una característica sexualment dimòrfica (diferent segons el sexe), tal com s'ha proposat per a Pteranodon.[2]
L'any 2006, Martill i Naish van trobar que les crestes de Tupuxuara i els seus parents es desenvolupaven per la part premaxil·lar de les crestes creixent cap enrere sobre el sostre del crani (tal com indica la sutura ben definida entre la premaxil·la i els ossos subjacents). El marge posterior de la part premaxil·lar de la cresta d'aquest exemplar només havia arribat per sobre del marge posterior de la fenestra nasoantorbitària, cosa que indica que no era un adult en el moment de la mort. Això suggereix que el desenvolupament de la cresta es va produir tard en el creixement d'un individu, probablement estava relacionat amb la visualització sexual i la maduresa sexual d'un determinat exemplar es podia avaluar per la mida i la disposició de la cresta. L'holotip de T. sethi, amb la seva cresta premaxil·lar hipertrofiada (engrandida), representaria així un individu adult vell (i l'estadi madur de Tupuxuara, segons la seva interpretació).[9] Kellner i Campos van trobar interessant la discussió de Martill i Naish sobre el desenvolupament de la cresta cranial, tot i que van trobar que el seu model proposat era especulatiu.[10]
-
 Vasos sanguinis a la cresta de l'holotip de T. sethi colat al CMNH
Vasos sanguinis a la cresta de l'holotip de T. sethi colat al CMNH -
 La cresta de T. sethi s'ha comparat amb el bec d'un tucà
La cresta de T. sethi s'ha comparat amb el bec d'un tucà
.jpg)

Els paleontòlegs David W. E. Hone, Naish i Innes C. Cuthill van reiterar la hipòtesi de creixement de Martill i Naish el 2012; atès que els pterosaures probablement eren precoços i podien volar poc després de l'eclosió, el paper de la cresta només era rellevant després de la maduresa (quan l'estructura estava totalment crescuda). Van considerar que la hipòtesi de la termoregulació era una explicació poc probable per als canals dels vasos sanguinis de la cresta, que van trobar compatibles amb l'alimentació del teixit en creixement (com la queratina del bec dels ocells). Hone, Naish i Cuthill van suggerir que les membranes de les ales i el sistema de sacs d'aire haurien estat més efectius per controlar la calor que una cresta, i el vent i l'aigua també podrien haver ajudat a refredar els pterosaures en entorns marítims d'alta temperatura.[21] El 2013, Witton va acceptar que les crestes substancialment més grans dels talassodromins adults indicaven que eren més importants per a les activitats de comportament que per a la fisiologia. Va trobar problemàtica la idea que les crestes s'utilitzaven per a la termoregulació, ja que no creixien regularment amb la mida del cos; van créixer a un ritme ràpid en gairebé adults, més ràpid del que es preveu per al creixement d'una estructura termoreguladora. Segons Witton, les membranes d'ala grans i altament vasculars dels pterosaures proporcionarien la superfície necessària per a la termoregulació, el que significa que les crestes no eren necessàries per a aquesta funció. Va concloure que els patrons dels vasos sanguinis de la cresta no difereixen gaire dels que es veuen als ossos sota els becs dels ocells, que s'utilitzen per transportar nutrients als ossos i teixits tous més que per a la termoregulació. Witton va assenyalar que, tot i que els becs dels ocells perden calor ràpidament, no es van desenvolupar per això; les crestes dels pterosaures també podrien haver tingut un efecte en la termoregulació, sense que aquesta fos la seva funció principal.[4]
Pêgas i els seus col·legues van assenyalar que s'ha proposat el dimorfisme sexual en la mida i la forma de la cresta per a alguns pterosaures; la forma de la cresta que es veu a l'holotip de T. sethi pot correlacionar amb un sexe i pot haver estat el resultat de la selecció sexual. Van suggerir que ambdós sexes podrien haver tingut crestes similars a causa de la selecció sexual mútua, però la interpretació de trets exagerats era difícil a causa de la petita mida de la mostra; caldria trobar més exemplars de T. sethi per avaluar aquestes teories. No van pensar que la termoregulació es correlacionés amb el creixement de la cresta en relació amb la mida del cos, ja que els becs dels tucans (el més gran dels ocells moderns) creixen dràsticament desproporcionat amb la mida corporal i funcionen com a estructures termoreguladores, a més de facilitar l'alimentació i el comportament social. Pêgas i els seus col·legues van trobar que l'estructura vascular dels becs del tucà era comparable a la de la cresta de T. sethi, i van concloure que la cresta també tenia múltiples funcions.[7]
Alimentació i dieta
[modifica]Kellner i Campos van trobar originalment les mandíbules de Thalassodromeus similars a les dels bec de tisora moderns (tres espècies d'ocells del gènere Rhynchops) amb les seves mandíbules comprimides lateralment, un bec semblant a una fulla i una mandíbula inferior que sobresurt (que s'assembla a les tisores a la vista lateral). Van argumentar que Thalassodromeus s'hauria alimentat d'una manera similar, tal com implica el nom del gènere; els bec de tisora passen per la superfície de l'aigua i submergeixen la mandíbula inferior per atrapar peixos i crustacis. Kellner i Campos van enumerar característiques addicionals del crani dels bec de tisora que són adaptacions per a l'alimentació piscívora, inclosos els ossos palatins engrandits, una característica també compartida amb Thalassodromeus. A diferència dels bec de tisora i altres pterosaures, els ossos palatins de Thalassodromeus eren còncaus, cosa que els escriptors van suggerir que podria haver ajudat a emmagatzemar aliments momentàniament. Igual que els bec de tisora, Thalassodromeus també sembla haver tingut músculs potents del coll, músculs de la mandíbula grans i una punta de la mandíbula superior ben irrigada per la sang (característiques que Kellner i Campos van interpretar com a adaptacions dels piscívors). Van concloure que el bec en forma de tisora i la cresta prima gairebé feien impossibles altres maneres de capturar preses, com ara baixar cap a l'aigua i submergir-s'hi. Concessant la dificultat de reconstruir el mètode de pesca de Thalassodromeus, el van imaginar amb un coll menys mòbil que els bec de tisora; amb la cresta impedint el seu cap de submergir-se, planejava, movent les ales només de tant en tant. Van trobar que el pterosaure amb mandíbules més semblants a les de Thalassodromeus era el més petit Rhamphorhynchus, tot i que creien que hauria tingut una capacitat limitada de pesca.[2][22]
-
 Originalment es va suggerir que T. sethi (dreta) s'alimentava com un bec de tisora existent (esquerra), però aquesta teoria ha estat criticada.
Originalment es va suggerir que T. sethi (dreta) s'alimentava com un bec de tisora existent (esquerra), però aquesta teoria ha estat criticada. -
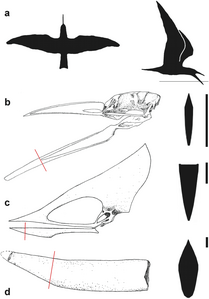 Comparació de la mandíbula d'un bec de tisora (a-b), Tupuxuara (c) i la punta de la mandíbula de T. oberlii (o Banguela, d), amb seccions transversals a la dreta.
Comparació de la mandíbula d'un bec de tisora (a-b), Tupuxuara (c) i la punta de la mandíbula de T. oberlii (o Banguela, d), amb seccions transversals a la dreta.


L'any 2004, el paleontòleg Sankar Chatterjee i l'enginyer R. Jack Templin van dir que els pterosaures més petits podrien haver pogut alimentar-se de peixos. Van dubtar que això fos possible per als més grans, a causa de la seva menor maniobrabilitat i capacitat de vol mentre es capbussaven. Chatterjee i Templin van assenyalar que els bec de tisora tenen el bec més contundent que els pterosaures com Thalassodromeus, per drenar l'aigua de la mandíbula mentre pesquen peixos.[23]
El 2007, el biofísic Stuart Humphries i els seus col·legues van qüestionar si algun pterosaure s'hauria alimentat habitualment de peixos i van dir que aquestes conclusions s'havien basat en comparacions anatòmiques més que en dades biomecàniques. Les proves de resistència que experimenten els becs d'ocells i les mandíbules de pterosaures es va provar hidrodinàmicament i aerodinàmicament mitjançant la creació d'un model d'un bec de tisora americà, d'un Thalassodromeus, i d'un Tupuxuara (que presumiblement no era piscívor), i remolcant-los al llarg d'un abeurador ple d'aigua a diferents velocitats. Els investigadors van trobar que els bec de tisora utilitzaven més energia per pescar del que es pensava anteriorment, i hauria estat impossible per a un pterosaure que pesés més d'1 kg pescar a causa de la potència metabòlica requerida. Van descobrir que fins i tot els pterosaures més petits, com Rhamphorhynchus, no estaven adaptats per a la pesca.[24][25] L'aparell d'alumini del model de Thalassodromeus es va destruir durant l'experiment, a causa de les forces altes i inestables que s'hi van exercir mentre pescava a gran velocitat, fet que va generar més dubtes sobre aquest mètode d'alimentació.[4] Els autors van utilitzar la punta de la mandíbula de T. oberlii per modelar el rendiment de Thalassodromeus, ja que en aquell moment va ser assignat a T. sethi.[7][12]
Unwin i Martill van suggerir l'any 2007 que els talassodromins podrien haver alimentat de manera similar a les cigonyes, com s'havia suggerit per als azhàrquids.[26] Witton va dir el 2013 que tot i que s'havia suggerit l'alimentació piscívora per a molts grups de pterosaures, la idea va ser criticada en els últims anys; els pterosaures no tenien pràcticament totes les adaptacions per a l'alimentació piscívora, per la qual cosa era poc probable que s'alimentessin d'aquesta manera. Thalassodromeus (a diferència dels bec de tisora) no tenia un crani especialment ample o robust o llocs d'adhesió als músculs de la mandíbula especialment grans, i la seva mandíbula era relativament curta i massissa. Witton estava d'acord amb Unwin i Martill que els talassodromins, amb les seves proporcions d'extremitats iguals i les mandíbules allargades, eren adequats per a caminar per la terra i alimentar-se de manera oportunista; els seus colls més curts i flexibles indicaven una manera diferent d'alimentar-se que els azhdarcoïdeus, que tenien colls més llargs i rígids. Va suggerir que els talassodromins podrien haver tingut hàbits d'alimentació més generalitzats, i els azhdarcoïdeus podrien haver estat més restringits; Thalassodromeus podria haver estat millor per caçar preses relativament grans i lluitadores que el seu parent, Tupuxuara, que tenia un crani més lleuger. Witton va subratllar que s'haurien de fer més estudis de morfologia funcional per il·luminar el tema i va especular que Thalassodromeus podria haver estat un depredador rapaç, utilitzant les seves mandíbules per sotmetre les preses amb mossegades fortes; el seu paladar còncau podria ajudar-lo a empassar-se grans preses.[27]
Pêgas i Kellner van presentar una reconstrucció dels músculs mandibulars de T. sethi en una conferència l'any 2015. Van trobar que els seus músculs de la mandíbula ben desenvolupats diferien dels del possible pescador amb immesió Anhanguera i dels azhdarcoïdeus que persegueixen animals terrestres, indicant que T. Sethi tenia una forta força de mossegada.[28] El 2018, Pêgas i els seus col·legues van acordar que les mandíbules robustes i semblants a una fulla de Thalassodromeus indicaven que les podria haver utilitzat per colpejar i matar preses, però van pensar que calia un treball biomecànic per corroborar la idea. Van trobar (a diferència de Witton) que Thalassodromeus tenia una articulació de la mandíbula reforçada i músculs robusts de la mandíbula, però calia més treball per determinar els seus hàbits dietètics. Segons Pêgas i els seus col·legues, l'articulació entre els ossos articulars i quadrats de T. sethi (on la mandíbula inferior connectada amb el crani) indica una obertura màxima de 50 graus, similar a la bretxa de 52 graus inferida per Quetzalcoatlus.[7]
Locomoció
[modifica]
En un comentari de 2002 sobre la descripció original de T. sethi, l'enginyer John Michael Williams va assenyalar que, tot i que Kellner i Campos havien esmentat que la gran cresta podria haver interferit aerodinàmicament durant el vol, no havien detallat aquest punt i havien comparat el pterosaure amb un ocell d'una cinquena part de la seva mida. Va suggerir que Thalassodromeus feia servir la seva cresta per equilibrar les seves mandíbules, amb el cap canviant d'actitud en funció del mode de locomoció. Williams va especular que la cresta seria inflable amb sang i presentava una resistència a l'aire variable, que va comparar amb un ventilador de mà; això hauria ajudat a l'animal a canviar l'actitud del cap durant el vol (i durant el contacte amb l'aigua), evitant que girés sense poderosos músculs del coll. La cresta hauria fet possibles vols llargs, en lloc d'interferir; Williams el va comparar amb l'espermaceti del cap del catxalot, afirmant que suposadament s'utilitza per canviar la flotabilitat mitjançant l'ajust de la temperatura. Kellner i Campos van rebutjar la idea d'una cresta inflable, ja que els seus ossos comprimits no ho permetrien; no van trobar convincent l'analogia del catxalot en relació als animals voladors, i van assenyalar que és més probable que l'espermaceti s'utilitzi durant l'agressió o per al sonar. Van acordar que la idea que la cresta tingués una funció en vol era temptadora i el moviment lateral del cap l'hauria ajudat a canviar de direcció, però s'haurien de fer estudis biomecànics i mecànics de vol de la cresta per determinar l'aerodinàmica de l'animal.[29]
Witton també va expressar l'esperança d'una anàlisi addicional de la locomoció dels talassodromins. Va assenyalar que com que les proporcions de les seves extremitats eren similars a les dels azhdarcoïdeus més ben estudiats, la forma de les seves ales i l'estil de vol podrien haver estat similars. Els talassodromins també podrien haver estat adaptats per al vol interior; les seves ales eren curtes i amples (a diferència de les ales llargues i estretes dels voladors marins), i eren més maniobrables i menys propensos a enganxar-se als obstacles. Sembla que els seus músculs inferiors de l'espatlla s'han engrandit, cosa que hauria ajudat amb potents (o freqüents) aletejos i capacitat d'enlairament. Encara que pot haver hagut de compensar la seva gran cresta durant el vol, el seu desenvolupament tardà en el creixement indica que no es va desenvolupar principalment per a l'aerodinàmica. Witton va suggerir que la similitud proporcional entre les extremitats dels talassodromins i els azhdarcoïdeus també indica que les seves habilitats terrestres haurien estat comparables. Les seves extremitats haurien estat capaces de fer llargs passos, i els seus peus curts i compactes haurien fet que aquesta mecànica fos eficient. Els músculs engrandits de les espatlles poden haver-los permès accelerar ràpidament quan corrien, i poden haver estat tan adaptats per al moviment a terra com s'ha suggerit per als azhàrquids; Witton va advertir que es necessitaven més anàlisis dels talassodromins per determinar-ho.[4]
Paleoecologia
[modifica]
Thalassodromeus es coneix de la Formació Romualdo, que data de l'etapa albià del període Cretaci inferior (fa uns 110 milions d'anys). La formació forma part del Grup Santana i, en el moment en què es va descriure Thalassodromeus, es pensava que era membre del que llavors es considerava la Formació Santana. La Formació Romualdo és una Lagerstätte (un jaciment sedimentari que conserva els fòssils en excel·lents condicions) que consisteix en concrecions de pedra calcària de la llacuna incrustades en esquists, i sobreposa la Formació Crato. És ben conegut per preservar fòssils tridimensionalment en concrecions calcàries, incloent molts fòssils de pterosaures. A més de fibres musculars de pterosaures i dinosaures, s'hi han trobat brànquies, tubs digestiu i cors que conserven peixos.[2][7] El clima tropical de la formació corresponia en gran part al clima brasiler actual.[30] La major part de la seva flora era xeròfita (adaptada a ambients secs). Les plantes més esteses van ser les Cycadales i la conífera Brachyphyllum.[31]
Altres pterosaures de la Formació Romualdo inclouen Anhanguera, Araripedactylus, Araripesaurus, Brasileodactylus, Cearadactylus, Coloborhynchus, Santanadactylus, Tapejara, Tupuxuara,[32] Barbosania,[33] Maaradactylus,[34] Tropeognathus[35] i Unwindia.[36] Els talassodromins només es coneixen d'aquesta formació, i tot i que s'han assignat restes postcranials ben conservades d'allà al grup, no es poden assignar al gènere a causa de la seva manca de cranis.[37][38] La fauna de dinosaures inclou teròpodes com Irritator, Santanaraptor, Mirischia i un dromeosaure indeterminat.[31] Pels jaciments es coneixen els crocodiliformes Araripesuchus i Caririsuchus,[39] així com les tortugues Brasilemys,[40] Cearachelys,[41] Araripemys, Euraxemys[42] i Santanachelys que també són coneguts dels dipòsits.[43] També hi havia concostracis, eriçons de mar, ostràcodes i mol·luscs.[30] Els fòssils de peixos ben conservats registren la presència de taurons hibodont, rinobàtids, lepisosteïformes, amiidae, pycnodontidae, aspidorhynchiformes, ichthyodectiformes, albúlids, cànnids, mawsoniidae i algunes formes incertes. Pêgas i els seus col·legues van assenyalar que els tàxons de pterosaures de la Formació Romualdo tenien diverses espècies: dues de Thalassodromeus, dues de Tupuxuara i fins a sis espècies d'Anhanguera. És possible que no totes les espècies de cada tàxon hagin coexistit en el temps (com s'ha proposat per als pteranodòntids de la Formació Niobrara), però no hi ha prou dades estratigràfiques perquè la Formació Romualdo ho comprovi.[7]
Referències
[modifica]- ↑ Entrada «Thalassodromeus» de la Paleobiology Database (en anglès). [Consulta: 3 juny 2023].
- ↑ 2,00 2,01 2,02 2,03 2,04 2,05 2,06 2,07 2,08 2,09 2,10 Kellner i Camos, 2002, p. 389-392.
- ↑ Veldmeijer, 2006, p. 11.
- ↑ 4,00 4,01 4,02 4,03 4,04 4,05 4,06 4,07 4,08 4,09 4,10 4,11 Witton, 2013, p. 234-243.
- ↑ Leonardi, 1984, p. 187-194.
- ↑ Kellner i Campos, 1990, p. 401-405.
- ↑ 7,00 7,01 7,02 7,03 7,04 7,05 7,06 7,07 7,08 7,09 7,10 7,11 7,12 7,13 7,14 7,15 Pêgas, Costa i Kellner, 2018, p. e1443273.
- ↑ 8,0 8,1 Veldmeijer, Signore i Meijer, 2005, p. 67-86.
- ↑ 9,0 9,1 9,2 Martill i Naish, 2006, p. 925-941.
- ↑ 10,0 10,1 10,2 10,3 Kellner i Campos, 2007, p. 1-14.
- ↑ 11,0 11,1 Witton, 2009, p. 1293-1300.
- ↑ 12,0 12,1 Headden i Campos, 2014, p. 815-826.
- ↑ Grellet Tinner i Codrea, 2015, p. 1673-1679.
- ↑ Dyke et al., Buffetaut, p. 1680-1682.
- ↑ Codrea i Grellet Tinner, 2015, p. 1683-1685.
- ↑ Witton, 2013, p. 51-52.
- ↑ 17,0 17,1 Kellner, 2004, p. 521-534.
- ↑ Unwin, 2003, p. 139-190.
- ↑ 19,0 19,1 Pinheiro et al., Bantim, p. 567-580.
- ↑ 20,0 20,1 Andres, Clark i Xu, 2014, p. 1011-1016.
- ↑ Hone, Naish i Cuthill, 2012, p. 139-156.
- ↑ Bogle, 2002.
- ↑ Chatterjee i Templin, 2004, p. 56.
- ↑ Humphries et al., 2007, p. e204.
- ↑ Gross, 2007, p. e217.
- ↑ Unwin i Martill, 2007, p. 475-524.
- ↑ Witton, 2013.
- ↑ Pêgas i kellner, 2015, p. 47-48.
- ↑ Williams, Kellner i Campos, 2002, p. 2207-2208.
- ↑ 30,0 30,1 Mabesoone i Tinoco, 1973, p. 97-118.
- ↑ 31,0 31,1 Martill et al., 2000, p. 891-900.
- ↑ Barrett et al., 2008, p. 61-107.
- ↑ Elgin i Frey, 2011, p. 259-275.
- ↑ Bantim et al., 2014, p. 201-223.
- ↑ Rodrigues i Kellner, 2013.
- ↑ Martill, 2011, p. 236-243.
- ↑ Aires et al., Pacheco, p. 343-355.
- ↑ Buchmann et al., 2017, p. 1139-1149.
- ↑ Figueiredo i Kellner, 2009, p. 323-331.
- ↑ de Lapparent de Broin, 2000, p. 43-95.
- ↑ Gaffney, Campos i Hirayama, 2001, p. 1-20.
- ↑ Gaffney, Tong i Meylan, 2009.
- ↑ Hirayama, 1998, p. 705-708.
Bibliografia
[modifica]- Aires, A. S. S.; Kellner, A. W. A.; Müller, R. T.; Da Silva, L. R.; Pacheco, C. P. «New postcranial elements of the Thalassodrominae (Pterodactyloidea, Tapejaridae) from the Romualdo Formation (Aptian-Albian), Santana Group, Araripe Basin, Brazil» (en anglès). Palaeontology, 57(2), 2014. DOI: 10.1111/pala.12069.
- Andres, B.; Clark, J.; Xu, X. «The Earliest Pterodactyloid and the Origin of the Group» (en anglès). Current Biology, 24(9), 2014, pàg. 1011-1016. DOI: 10.1016/j.cub.2014.03.030. PMID: 24768054.
- Bantim, R. A. M.; Saraiva, A. A. F.; Oliveira, G. R.; Sayão, J. M. «A new toothed pterosaur (Pterodactyloidea: Anhangueridae) from the Early Cretaceous Romualdo Formation, NE Brazil» (en anglès). Zootaxa, 3869(3), 2014. DOI: 10.11646/zootaxa.3869.3.1. PMID: 25283914.
- Barrett, P.; Butler, R.; Edwards, N.; Milner, A. R. «Pterosaur distribution in time and space: An atlas» (en anglès). Zitteliana Reihe B: Abhandlungen der Bayerischen Staatssammlung für Paläontologie und Geologie, 28, 2008.
- Bogle, L. S. «New find: pterosaur had strange crest, fishing style» (en anglès). National Geographic, 2002. Arxivat de l'original el 2017-01-26 [Consulta: 8 desembre 2023].
- Brito, P.; Yabumoto, Y. «An updated review of the fish faunas from the Crato and Santana formations in Brazil, a close relationship to the Tethys fauna» (en anglès). Bulletin of Kitakyushu Museum of Natural History and Human History, 9, 2011.
- Buchmann, R.; Rodrigues, T.; Polegario, S.; Kellner, A. W. A. «New information on the postcranial skeleton of the Thalassodrominae (Pterosauria, Pterodactyloidea, Tapejaridae)» (en anglès). Historical Biology, 30(8), 2017. DOI: 10.1080/08912963.2017.1343314.
- Chatterjee, S.; Templin, R. J.. {{{títol}}} (en anglès). 376, 2004. DOI 10.1130/0-8137-2376-0.1. ISBN 9780813723761.
- Codrea, V. A.; Grellet Tinner, G. «Reply to comment by Dyke et al. on "Thalassodromeus sebesensis, an out of place and out of time Gondwanan tapejarid pterosaur" by Grellet-Tinner and Codrea» (en anglès). Gondwana Research, 27(4), 2015. Bibcode: 2015GondR..27.1683C. DOI: 10.1016/j.gr.2014.08.003.
- de Lapparent de Broin, F. «The oldest pre-podocnemidid turtle (Chelonii, Pleurodira), from the early Cretaceous, Ceará state, Brasil, and its environment» (en anglès). Treballs del Museu de Geologia de Barcelona, 9, 2000. ISSN: 2385-4499.
- Dyke, G.; Vremir, M.; Brusatte, S. L.; Bever, G. S.; Buffetaut, E. «Thalassodromeus sebesensis - A new name for an old turtle. Comment on "Thalassodromeus sebesensis, an out of place and out of time Gondwanan tapejarid pterosaur", Grellet-Tinner and» (
 PDF) (en anglès). Gondwana Research, 27(4), 2015. Bibcode: 2015GondR..27.1680D. DOI: 10.1016/j.gr.2014.08.004.
PDF) (en anglès). Gondwana Research, 27(4), 2015. Bibcode: 2015GondR..27.1680D. DOI: 10.1016/j.gr.2014.08.004. - Elgin, A.; Frey, E. «A new ornithocheirid, Barbosania gracilirostris gen. et sp. nov. (Pterosauria, Pterodactyloidea) from the Santana Formation (Cretaceous) of NE Brazil» (en anglès). Swiss Journal of Palaeontology, 130(2), 2011. DOI: 10.1007/s13358-011-0017-4.
- Figueiredo, R. G.; Kellner, A. W. A. «A new crocodylomorph specimen from the Araripe Basin (Crato Member, Santana Formation), northeastern Brazil» (en anglès). Paläontologische Zeitschrift, 83(2), 2009. DOI: 10.1007/s12542-009-0016-6.
- Gaffney, E. S.; Campos, D. A.; Hirayama, R. «Cearachelys, a new side-necked turtle (Pelomedusoides: Bothremydidae) from the Early Cretaceous of Brazil» (en anglès). American Museum Novitates, 3319, 2001. DOI: 10.1206/0003-0082(2001)319<0001:CANSNT>2.0.CO;2.
- Gaffney, E. S.; Tong, H.; Meylan, P. A. «Evolution of the side-necked turtles: The families Bothremydidae, Euraxemydidae, and Araripemydidae» (en anglès). Bulletin of the American Museum of Natural History, 300, 2009. DOI: 10.1206/0003-0090(2006)300[1:EOTSTT]2.0.CO;2.
- Grellet Tinner, G.; Codrea, V. A. «Thalassodromeus sebesensis, an out of place and out of time Gondwanan tapejarid pterosaur» (en anglès). Gondwana Research, 27(4), 2015. Bibcode: 2015GondR..27.1673G. DOI: 10.1016/j.gr.2014.06.002.
- Gross, L. «A new view of pterosaur feeding habits» (en anglès). PLOS Biology, 5(8), 2007. DOI: 10.1371/journal.pbio.0050217. PMC: 1925131. PMID: 20076682.
- Headden, J. A.; Campos, H. B. N. «An unusual edentulous pterosaur from the Early Cretaceous Romualdo Formation of Brazil» (en anglès). Historical Biology, 27(7), 2014. DOI: 10.1080/08912963.2014.904302.
- Hirayama, R. «Oldest known sea turtle» (en anglès). Nature, 392(6677), 1998. Bibcode: 1998Natur.392..705H. DOI: 10.1038/33669. ISSN: 0028-0836.
- Hone, D. W. E.; Naish, D.; Cuthill, I. C. «Does mutual sexual selection explain the evolution of head crests in pterosaurs and dinosaurs?» (en anglès). Lethaia, 45(2), 2012. DOI: 10.1111/j.1502-3931.2011.00300.x.
- Humphries, S.; Bonser, R. H. C.; Witton, M. P.; Martill, D. M. «Did pterosaurs feed by skimming? Physical modelling and anatomical evaluation of an unusual feeding method» (en anglès). PLOS Biology, 5(8), 2007. DOI: 10.1371/journal.pbio.0050204. PMC: 1925135. PMID: 17676976.
- Kellner, A. W. A.; Campos, D. A. «Preliminary description of an unusual pterosaur skull of the Lower Cretaceous from the Araripe Basin» (en anglès). Atas - Simposio Sobre a Bacia do Araripe e Bacias Interiores do Nordeste, 1990.
- Kellner, A. W. A.; Campos, D. A. «The function of the cranial crest and jaws of a unique pterosaur from the early Cretaceous of Brazil» (en anglès). Science, 297(5580), 2002. Bibcode: 2002Sci...297..389K. DOI: 10.1126/science.1073186. PMID: 12130783.
- Kellner, A. W. A. «New information on the Tapejaridae (Pterosauria, Pterodactyloidea) and discussion of the relationships of this clade» (en anglès). Ameghiniana, 41(4), 2004. ISSN: 1851-8044.
- Kellner, A. W. A.; Campos, D. A. «Short note on the ingroup relationships of the Tapejaridae (Pterosauria, Pterodactyloidea)» (en anglès). Boletim do Museu Nacional: Geologia, 75, 2007.
- Leonardi, G. «I rettili volanti». A: Sulle Orme Dei Dinosauri (en italià). Venècia: Erizzo Editrice, 1984. ISBN 978-8870770186.
- Mabesoone, J. M.; Tinoco, I. M. «Palaeoecology of the Aptian Santana Formation (Northeastern Brazil)» (en anglès). Palaeogeography, Palaeoclimatology, Palaeoecology, 14(2), 1973, pàg. 97-118. Bibcode: 1973PPP....14...97M. DOI: 10.1016/0031-0182(73)90006-0. ISSN: 0031-0182.
- Martill, D.; Frey, E.; Sues, H. D.; Cruickshank, A. «Skeletal remains of a small theropod dinosaur with associated soft structures from the Lower Cretaceous Santana Formation of NE Brazil» (en anglès). Canadian Journal of Earth Sciences, 37(6), 2000. Bibcode: 2000CaJES..37..891M. DOI: 10.1139/cjes-37-6-891.
- Martill, D. M.; Naish, D. «Cranial crest development in the Azhdarchoid pterosaur Tupuxuara, with a review of the genus and tapejarid monophyly» (en anglès). Palaeontology, 49(4), 2006. DOI: 10.1111/j.1475-4983.2006.00575.x.
- Martill, D. M. «A new pterodactyloid pterosaur from the Santana Formation (Cretaceous) of Brazil» (en anglès). Cretaceous Research, 32(2), 2011. DOI: 10.1016/j.cretres.2010.12.008.
- Pêgas, R. V.; Kellner, A. W. A. «Preliminary mandibular myological reconstruction of Thalassodromeus sethi (Pterodactyloidea: Tapejaridae)» (en anglès). Flugsaurier 2015 Portsmouth, Abstracts, 2015.
- Pêgas, R. V.; Costa, F. R.; Kellner, A. W. A. «New Information on the osteology and a taxonomic revision of The genus Thalassodromeus (Pterodactyloidea, Tapejaridae, Thalassodrominae)» (en anglès). Journal of Vertebrate Paleontology, 38 (2), 2018. DOI: 10.1080/02724634.2018.1443273.
- Pinheiro, F. L.; Fortier, D. C.; Schultz, C. L.; De Andrade, J. A. F. G.; Bantim, R. A. M. «New information on Tupandactylus imperator, with comments on the relationships of Tapejaridae (Pterosauria)» ( PDF) (en anglès). Acta Palaeontologica Polonica, 56(3), 2011. DOI: 10.4202/app.2010.0057.
- Rodrigues, T.; Kellner, A. W. A. «Taxonomic review of the Ornithocheirus complex (Pterosauria) from the Cretaceous of England» (en anglès). ZooKeys, 308, 2013. DOI: 10.3897/zookeys.308.5559. PMC: 3689139. PMID: 23794925.
- Unwin, D. M. «On the phylogeny and evolutionary history of pterosaurs» (en anglès). Geological Society, London, Special Publications, 217(1), 2003. Bibcode: 2003GSLSP.217..139U. DOI: 10.1144/GSL.SP.2003.217.01.11.
- Unwin, D.; Martill, D. «Pterosaurs of the Crato Formation». A: The Crato Fossil Beds of Brazil: Window into an Ancient World (en anglès). Cambridge: Cambridge University Press, 2007. ISBN 9780521858670.
- Veldmeijer, A. J.; Signore, M.; Meijer, H. J. M. «Description of two pterosaur (Pterodactyloidea) mandibles from the upper Cretaceous Santana Formation, Brazil» (en anglès). Deinsea, 11, 2005.
- Veldmeijer, A. J.. Toothed pterosaurs from the Santana Formation (Cretaceous; Aptian-Albian) of northeastern Brazil (en anglès). Proefschrift Universiteit Utrecht, 2006.
- Williams, J. M.; Kellner, A. W. A.; Campos, D. A. «Form, function, and the flight of the pterosaur» (en anglès). Science, 297(5590), 2002. DOI: 10.1126/science.297.5590.2207b. JSTOR: 3832354. PMID: 12353519.
- Witton, M. P. «A new species of Tupuxuara (Thalassodromidae, Azhdarchoidea) from the Lower Cretaceous Santana Formation of Brazil, with a note on the nomenclature of Thalassodromidae» (en anglès). Cretaceous Research, 30(5), 2009. DOI: 10.1016/j.cretres.2009.07.006.
- Witton, M. P.. Pterosaurs: Natural History, Evolution, Anatomy (en anglès). Princeton; Oxford: Princeton University Press, 2013. ISBN 978-0-691-15061-1.












